

 LINKED PAPER
LINKED PAPER
Quantifying inter-annual variability on the space-use of parental Northern Gannets (Morus bassanus) in pursuit of different prey types. d’Entremont, K.J.N, Pratte, I., Gjerdrum, C., Wong, S.N.P. & Montevecchi, W.A. 2023 PLoS One. doi: 10.1371/journal.pone.0288650 VIEW
Examination of the foraging behaviour (e.g., trip distance, number of dives, space-use, etc.) of marine birds can provide considerable information on the distribution of their pelagic prey and how environmental conditions impact their spatial and temporal availability. Northern Gannets (Morus bassanus; hereafter gannets) are generalist foragers, which typically exhibit intra-annual shifts in foraging tactics based on the availability of their spatio-temporally constrained prey (e.g., d’Entremont et al. 2022). For example, gannets exhibit two different dive profiles, based on the prey type they are targeting: deep, prolonged, wing-propelled, “U-shaped” dives, and rapid, shallow, “V-shaped” dives (for example, Garthe et al. 2000).
In the Northwest Atlantic, U-shaped dives by gannets are almost exclusively associated with the pursuit and capture of Capelin (Mallotus villosus), often capturing several fish per dive. Gannets use V-shaped dives in pursuit of various, larger, faster moving pelagic fishes such as Atlantic Mackerel (Scomber scombrus) Atlantic Herring (Clupea harengus) and Atlantic Saury (Scomberesox saurus), typically capturing only one individual per dive. The availability and spatio-temporal distribution of these prey species varies intra-annually with Capelin preferring colder temperatures (<12°C) for inshore spawning in June and July, Atlantic Herring preferring temperatures of 8-12°C in May, June and July, Atlantic Mackerel migrating through the region in August and September when their preferred temperature range of 9-13°C occurs, and Atlantic Saury being a late migrant with preferred temperatures of ~18°C (Dudley et al. 1985, Brennan et al. 2016, Crook et al. 2017, Olafsdottir et al. 2019).
With Capelin being a critical prey species for gannets in early chick-rearing and Atlantic Mackeral/Atlantic Saury being key in late chick-rearing, we aimed to create habitat suitability models using GPS locations from gannets associated with each dive profile to inform us on how their space-use differs when in pursuit of different prey species. These models also provide critical information on suitable conditions for the occurrence of their pelagic prey. From 2019 to 2021, we deployed 25 GPS/Time-depth recorder devices on parental Northern Gannets during chick-rearing (July to October). Using location and dive data derived from these devices, we created three separate habitat suitability models using all U-shaped dive locations, V-shaped dive locations in early chick-rearing and V-shaped dive locations in late chick-rearing. Covariates examined in our analysis included sea surface temperature (SST), bathymetric depth, slope, distance to colony, longitude and latitude.
We found that space-use by gannets when foraging for Capelin in early chick-rearing (e.g., U-shaped dives), was typically confined to coastal areas. Meanwhile, space-use when employing V-shaped dives varied intra-annually, with a preference for shallow coastal waters in early chick-rearing (e.g., Atlantic Herring, Atlantic Mackerel and possibly Capelin occurring near the surface), and a preference for shallow waters as well as deeper pelagic waters in late chick-rearing (e.g., Atlantic Mackerel and Atlantic Saury). We show that parental gannets are susceptible to different anthropogenic risks depending on time of year and the prey species they are in pursuit of while foraging. When Capelin is spawning in the region during early chick-rearing, gannets are most at risk of interaction with inshore fisheries while they pursue this prey base in coastal waters. Meanwhile, gannets are more at risk to interact with offshore vessels such as oil tankers during late chick-rearing when in pursuit of Atlantic Mackerel and Atlantic Saury.
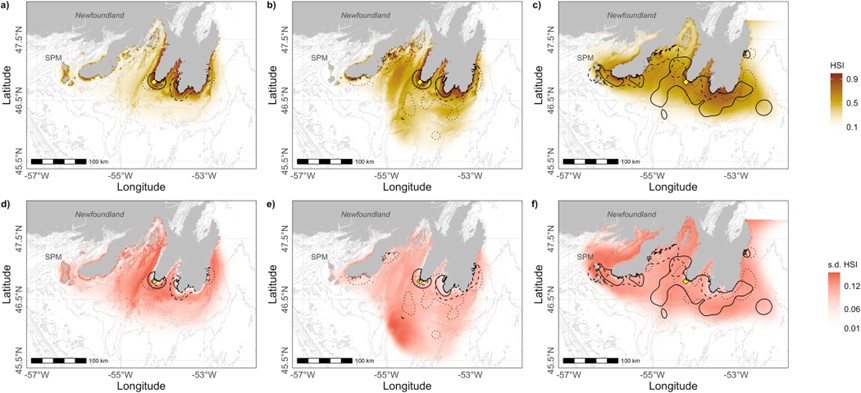
Figure 1 Spatial habitat predictions for a) U-shaped dives, b) early and c) late V-shaped dives of Northern Gannets tracked from Cape St. Mary’s (yellow diamond) during three breeding seasons. Average of daily habitat suitability index (HSI) scaled from 0 (unsuitable) to 1 (highly suitable) (a, b, c), and standard deviation of daily HSI for d) U-shaped dives, e) early and f) late V-shaped dives. The spatial resolution of the prediction was set to the lowest resolution of our environmental layers (SST, at 1000m). Utilization distributions at 50% for each dive type are overlaid on the top panels for each year represented by different line types (solid—2019, dashed—2020, dotted—2021).
Habitat models also revealed that suitable habitat for U-shaped dives (e.g., the pursuit of Capelin) included flat bottom relief with SST of 11-15°C. Suitable habitat for V-dives included steeper slopes and shallow waters with SST of 12-15°C and ~18°C during early chick-rearing, with suitable habitat for late chick-rearing V-shaped dives including shallow coastal waters and pelagic waters, with SST >16°C. The variability observed in the V-shaped dive models likely reflects the variation of thermal preferences by prey species pursued when employing this dive profile.
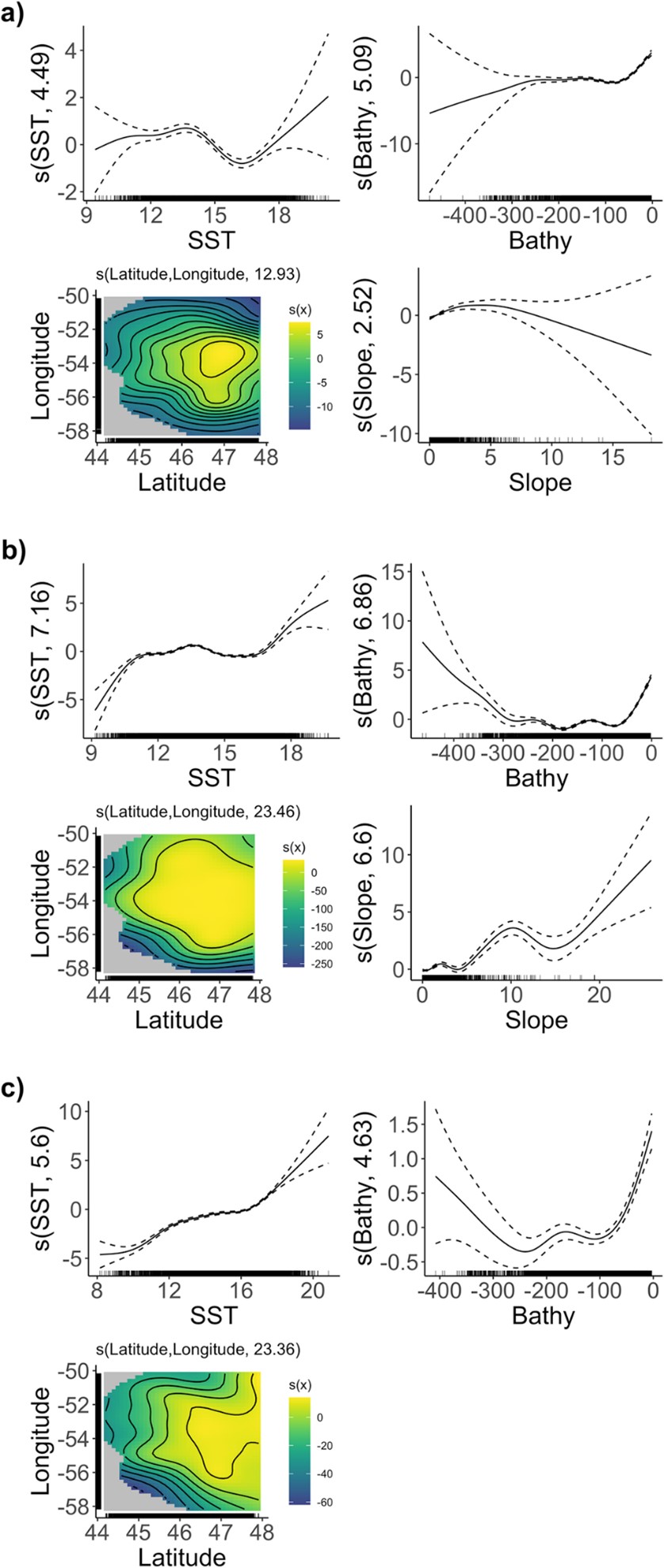
Figure 2 Model variables for estimated probability of occurrence of a) U-shaped dives and b) early and c) late V-shaped dives by Northern Gannets tracked during three breeding seasons (2019–2021) at the colony of Cape St. Mary’s, Newfoundland. The y-axis represents the function of each predictor variable with the effective degrees of freedom (edf) of the smooth term in brackets. 0 on the y-axis corresponds to absence of an effect of the predictor variable on the estimated probability of occurrence. The confidence intervals (dashed lines) are shown at 1 standard error above and below the smoothed estimate.
These habitat suitability models created using dive locations associated with different prey types can also be used as proxies for determining suitable habitat for the prey that gannets are exploiting with different foraging tactics. In particular, the U-shaped dive model is indicative of suitable habitat/habitat requirements for Capelin, a keystone prey species for several top predators (e.g., Common Murre (Uria aalge), Atlantic Puffin (Fratercula arctica), Atlantic Cod (Gadus morhua)) within the Northwest Atlantic ecosystem. As such, areas deemed as highly suitable for the pursuit of Capelin by gannets in our study likely represent areas of multi-species management concern. The V-shaped dive model can also inform us on habitat requirements of prey species such as Atlantic Mackerel and Atlantic Saury, though not as refined as the U-shaped model owing to lack of species specificity. Thus, we suggest that habitat suitability of organisms employing different foraging tactics for different prey species is a valuable tool for determining how space-use may differ when they are reliant on temporally and environmentally constrained prey sources.
References
Brennan, C.E., Blanchard, H. & Fennel, K. 2016. Putting temperature and oxygen thresholds of marine animals in context of environmental change: A regional perspective for the Scotian Shelf and Gulf of St. Lawrence. PLoS One 11:e0167411. VIEW
Crook, K.A., Maxner, E. & Davoren, G.K. 2017. Temperature-based spawning habitat selection by capelin (Mallotus villosus) in Newfoundland. ICES Journal of Marine Science 74:1622-1629. VIEW
d’Entremont, K.J.N., Davoren, G.K., Walsh, C.J., Wilhelm, S.I. & Montevecchi, W.A. 2022. Intra- and inter-annual shifts in foraging tactics by parental northern gannets Morus bassanus indicate changing prey fields. Marine Ecology Progress Series 698:155-170. VIEW
Dudley, S.F.J., Field, J.G. & Shelton, P.A. 1985. Distribution and abundance of eggs, larvae and early juveniles of saury Scomberesox saurus scombroides (Richardson) off the south-western Cape, South Africa, 1977/78. South African Journal of Marine Science 3:229-237. VIEW
Garthe, S., Benvenuti, S. & Montevecchi, W.A. 2000. Pursuit plunging by northern gannets (Sula bassana) feeding on capelin (Mallotus villosus). Proceedings of the Royal Society B 267:1717-1722. VIEW
Olafsdottir, A.H., Utne, K.R., Jacobsen, J.A., Jansen, T., Óskarsson, G.J., Nøttestad, L., et al. 2019. Geographical expansion of Northeast Atlantic mackerel (Scomber scombrus) in the Nordic Seas from 2007 to 2016 was primarily driven by stock size and constrained by low temperatures. Deep Sea Research Part II: Topical Studies in Oceanography 159:152-168. VIEW
Image credit
Top right: © Megan Buers.
If you want to write about your research in #theBOUblog, then please see here.

