

LINKED PAPER
Seasonal metabolic adjustments in an avian evolutionary relict restricted to mountain habitat. Oswald, K.N., Lee, A.T.K, Smit, B. 2021 Journal of Thermal Biology. DOI: j.jtherbio.2020.102815 VIEW
 When we set out to measure cold tolerance in Cape Rockjumpers (Chaetops frenatus) our question seemed relatively simple: did living in the relatively mild climate of the mountain Fynbos mean Rockjumpers had physiological specializations for cooler conditions? Cape Rockjumpers are a great species to study as they are a range-restricted mountain bird that remains in the cool mountain-tops of the Cape Fold Mountains year-round. They also make for an interesting case study into early passerine metabolism. Cape Rockjumpers are one of two species in the genus Chaetops, the sole genus within the family Chaetopidae. They are part of an early radiation of oscine passerines into Africa, diverging from other passerines ~21 million years ago (Oliveros et al. 2019).
When we set out to measure cold tolerance in Cape Rockjumpers (Chaetops frenatus) our question seemed relatively simple: did living in the relatively mild climate of the mountain Fynbos mean Rockjumpers had physiological specializations for cooler conditions? Cape Rockjumpers are a great species to study as they are a range-restricted mountain bird that remains in the cool mountain-tops of the Cape Fold Mountains year-round. They also make for an interesting case study into early passerine metabolism. Cape Rockjumpers are one of two species in the genus Chaetops, the sole genus within the family Chaetopidae. They are part of an early radiation of oscine passerines into Africa, diverging from other passerines ~21 million years ago (Oliveros et al. 2019).
In colder temperatures, birds will allocate a large amount of energy toward staying warm. Cold tolerance is generally measured using (1) a minimum metabolic rate, (2) a maximum metabolic rate, and (3) the metabolic breadth between their lowest and highest metabolic rates (metabolic expansibility).
Comparing these responses between winter and summer can be used to give us an idea of how much flexibility is present within a species. However, physiological traits not only vary between season, but within and among populations (Noakes et al. 2017; Noakes and McKechnie 2020), with altitude (Cooper 2002), with latitude (McKechnie et al. 2015), and with phylogeny (Swanson and Bozinovic 2011).
 Figure 1 Cape Rockjumpers (male pictured here) were caught using baited snap-traps, and then transported to a main field station at Blue Hill Nature Reserve 1–5 km from the capture site. They were kept between 18–48 hours before being released back at the site of capture.
Figure 1 Cape Rockjumpers (male pictured here) were caught using baited snap-traps, and then transported to a main field station at Blue Hill Nature Reserve 1–5 km from the capture site. They were kept between 18–48 hours before being released back at the site of capture.
We captured 12 non-breeding adults in July-August 2015 (summer) and 12 again in February 2016 (winter), at Blue Hill Nature Reserve in the Western Cape of South Africa. We brought them to a field station set up at Blue Hill Nature Reserve and kept them for either two nights (the first 5 individuals in each season, to determine baseline data) or one night in order to collect our physiological measurements.
 Figure 2 A male Cape Rockjumper puffs out his feathers as the temperature in the chamber is decreased at a constant rate. As the temperature drops, he raises his metabolic heat production to stay warm. We used HelOx as this allowed maximum heat loss to be achieved at warmer temperatures than would occur in normal air, and so the bird faces much lower risk.
Figure 2 A male Cape Rockjumper puffs out his feathers as the temperature in the chamber is decreased at a constant rate. As the temperature drops, he raises his metabolic heat production to stay warm. We used HelOx as this allowed maximum heat loss to be achieved at warmer temperatures than would occur in normal air, and so the bird faces much lower risk.
In winter, Cape Rockjumpers were in better body condition and seemed to have metabolic responses centered on conserving energy –– winter birds had a lower minimum metabolic rate, lower body temperature, and better insulation, all of which would help reduce energy expenditure. They also had a greater ability to increase their maximum metabolic rate, which would come in handy to stay warm during the coldest days.
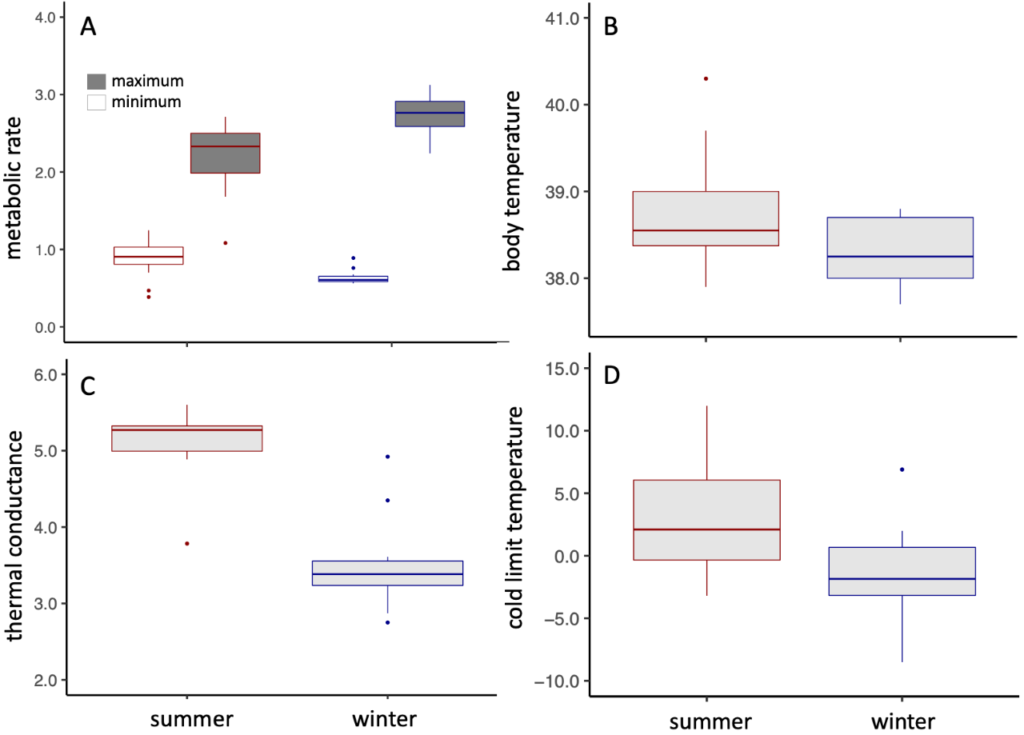 Figure 3 Seasonal metabolic data (blue = winter, red = summer) including A) minimum metabolic rate (Watts) and maximum metabolic rate (Watts), B) body temperature (°C), C) thermal conductance (J g−1 h−1 °C−1), and D) cold limit temperature (the HelOx temperature at which birds reached maximum metabolic rate; °C). Figure was modified from Oswald et al. 2021.
Figure 3 Seasonal metabolic data (blue = winter, red = summer) including A) minimum metabolic rate (Watts) and maximum metabolic rate (Watts), B) body temperature (°C), C) thermal conductance (J g−1 h−1 °C−1), and D) cold limit temperature (the HelOx temperature at which birds reached maximum metabolic rate; °C). Figure was modified from Oswald et al. 2021.
Cape Rockjumpers continue to present a bit of an enigma. For example, why were winter birds in better condition than summer birds despite food availability being highest in spring/early summer? This could be due to birds facing higher energy demands in summer, when they would be at the end of a long and demanding breeding season (August to December). Cape Rockjumpers also had very low values of metabolic expansibility, but these are notoriously difficult to interpret (Noakes et al. 2017; Noakes and McKechnie 2020).
Still, while Cape Rockjumper physiological traits seem to contrast to other subtropical passerines it is hard to differentiate between traits that are used for improved cold tolerance, and those that are reflective of phylogeny. Our values in general seemed quite low compared to those of other passerines, which made us question whether Cape Rockjumper physiological data may be reflective of their basal placement in passerine phylogeny. After doing a phylogenetic analysis, fitting Cape Rockjumper data into available data published for other passerines, Cape Rockjumpers stood out for having metabolic patterns that seemed to fit better with suboscines (earlier passerines) compared to oscines (later passerines). Cape Rockjumpers are also similar to suboscines in their slower life history (Ricklefs and Wikelski 2002) and so their physiological patterns may reflect a difference in pace of life; of course, this itself could also be related to phylogeny.
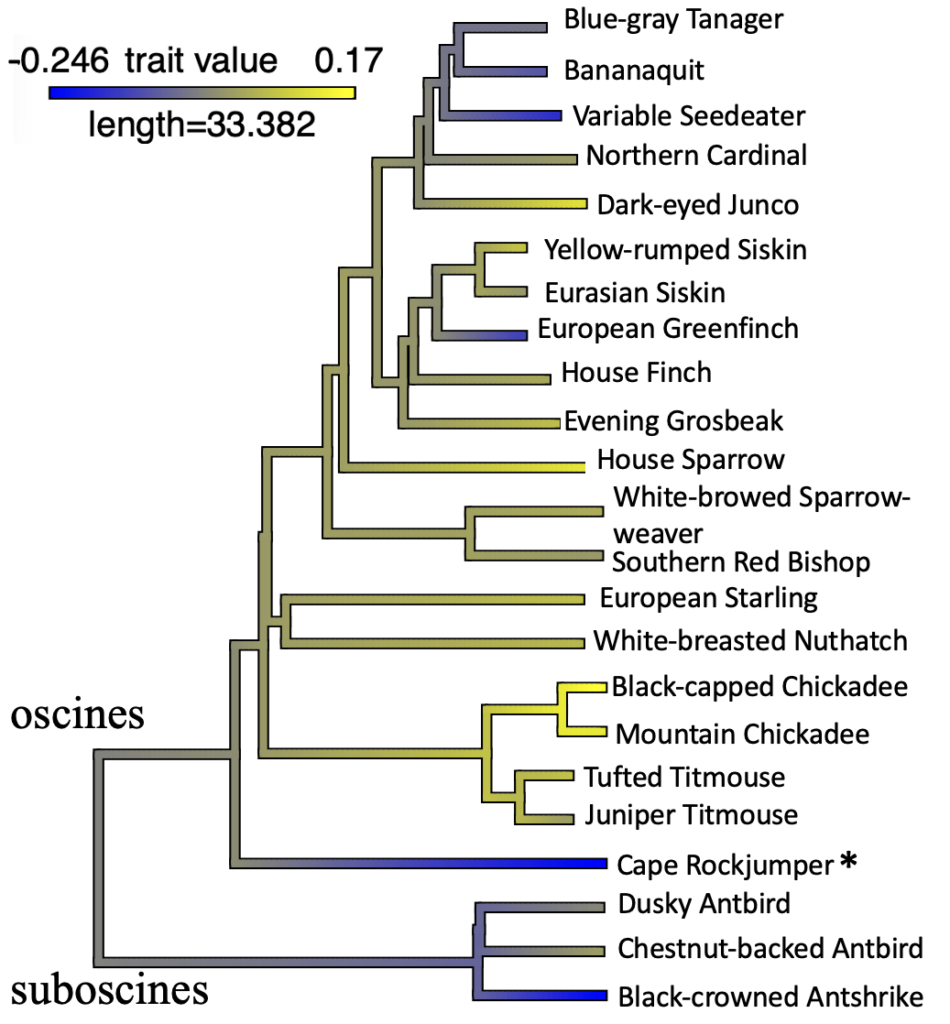 Figure 4 Phylogeny of residual values for expected log10-transformed winter maximum thermogenic capacity (Msum; Watts) data of passerine birds in winter (n = 23). Data for Cape Rockjumpers (Chaetops frenatus) were added to existing data. Asterisk indicates non-significant Bonferroni adjusted outliers. Figure was modified from Oswald et al. 2021, species names are consistent with Birds of the World.
Figure 4 Phylogeny of residual values for expected log10-transformed winter maximum thermogenic capacity (Msum; Watts) data of passerine birds in winter (n = 23). Data for Cape Rockjumpers (Chaetops frenatus) were added to existing data. Asterisk indicates non-significant Bonferroni adjusted outliers. Figure was modified from Oswald et al. 2021, species names are consistent with Birds of the World.
Cape Rockjumper metabolic traits do not seem to fit the “normal” patterns often found in other South African passerines (McKechnie et al. 2015; Stager et al. 2016), and so our results add to the growing evidence that metabolic responses are diverse at subtropical latitudes. In addition, Cape Rockjumpers’ summit metabolism and metabolic expansibility seemed more reflective of suboscines as opposed to oscines. Our study further highlights the need to examine physiological responses not only from different seasons, but also from diverse geographic areas and diverse phylogenetic lineages.
References
Oliveros, C.H., Field, D.J., Ksepka, D.T., Barker, F.K., Aleixo, A., Andersen, M.J., Alström, P., Benz, B.W., Braun, E.L., Braun, M.J., Bravo, G.A., Brumfield, R.T., Chesser, R.T., Claramunt, S., Cracraft, J., Cuervo, A.M., Derryberry, E.P., Glenn, T.C., Harvey, M.G., Hosner, P.A., Joseph, L., Kimball, R.T., Mack, A.L., Miskelly, C.M., Peterson, A.T., Robbins, M.B., Sheldon, F.H., Silveira, L.F., Tilston Smith, B., White, N.D., Moyle, R.G., & Faircloth, B.C. 2019. Earth history and the passerine superradiation. Proceedings of the National Academy of Sciences 116: 7916-7925. VIEW
Noakes, M.J., Wolf, B.O. & McKechnie, A.E. 2017. Seasonal metabolic acclimatization varies in direction and magnitude among populations of an Afrotropical passerine bird. Physiological and Biochemical Zoology 90: 178-189. VIEW
Noakes, M.J. & McKechnie, A.E. 2020. Seasonal metabolic acclimatization varies in direction and magnitude among years in two arid-zone passerines. Physiological and Biochemical Zoology 93: 140-152. VIEW
Cooper, S.J. 2002. Seasonal metabolic acclimatization in mountain chickadees and juniper titmice. Physiological and Biochemical Zoology 75: 386-395. VIEW
McKechnie, A.E., Noakes, M.J. & Smit, B. 2015. Global patterns of seasonal acclimatization in avian resting metabolic rates. Journal of Ornithology 156: 367-376. VIEW
Swanson, D.L. & Bozinovic, F. 2011. Metabolic capacity and the evolution of biogeographic patterns in oscine and suboscine passerine birds. Physiological and Biochemical Zoology 84: 185-194. VIEW
Ricklefs, R.E. & Wikelski, M. 2002. The physiology/life-history nexus. Trends in Ecology and Evolution 72: 462-468. VIEW
Image credit
Top right: Male Cape Rockjumper at Blue Hill Nature Reserve in South Africa. © Krista Oswald

