


Kittiwake © Wesley Davies
Where do seabirds forage? And why do we need to know? One population of Kittiwakes may provide some of the answers
LINKED PAPER
Inter- and intra-year variation in foraging areas
of breeding Kittiwakes (Rissa tridactyla). Robertson, G.S., Bolton, M., Grecian, W.J. & Monaghan, P. 2014. Marine Biology 161: 1973–1986. DOI: 10.1007/s00227-014-2477-8
Seabird populations around the world have declined in recent years. These declines have been linked to human activities such as offshore development, fishery bycatch and pollution; however, one of the main factors influencing changes in seabird abundance and breeding success has been a reduction in food availability (Oro and Furness 2002, Cury et al. 2011). Seabirds feed on a variety of different prey types and sizes, and reliance on particular prey types can vary among regions. Seabirds in the North Sea rely heavily on Lesser Sandeel Ammodytes marinus during the breeding season, and declines in seabird populations in the North Sea have been linked to reductions in the abundance of sandeel (Rindorf et al. 2000).
Seabird species vary in vulnerability to sandeel declines around breeding colonies (Furness and Tasker 2000). Diving species, such as guillemots and razorbills, which have varied diets and are able to forage for food deep in the water column, are less affected by years of low sandeel abundance than surface-feeding species. The breeding success of small surface-feeders has been shown to be highly dependent on annual sandeel abundance during the breeding season (Lewis et al. 2001, Frederiksen et al. 2004). Kittiwakes and terns forage close to the water surface and rely on prey being available within range of their limited diving abilities. Climate change and the activities of industrial fisheries have reduced the abundance and availability of sandeel to breeding seabirds since the 1980s, with corresponding declines in Kittiwake and tern breeding success (Monaghan 1992, Frederiksen et al. 2004).
While legal protection is afforded to seabird breeding colonies, little protection is currently given to offshore foraging areas. It is becoming evident that important foraging areas need to be protected if recent declines in the breeding success of vulnerable seabird species are to be reversed. The UK Government is a signatory to international agreements including the EU Birds Directive, Convention on Biological Diversity and the OSPAR Convention whose aims include establishing a network of Marine Protected Areas (MPAs) and Special Protection Areas incorporating foraging areas used by seabirds. The size and location of important seabird foraging areas must be determined for MPAs to be effective. The recent miniaturisation of Global Positioning System (GPS) devices has allowed seabirds to be tracked to and from their foraging grounds and for foraging areas of a variety of seabird species to be identified. Tracking data can also be used to identify environmental variables associated with foraging locations, and for probable foraging areas to be determined over wider areas.
Until recently, many tracking studies have been carried out during a single breeding season (Kotzerka et al. 2010, Votier et al. 2010), or breeding stage (Stauss et al. 2012, Chivers et al. 2013), and as such represent foraging behaviour over a restricted temporal scale. Little is known about the variability of species’ foraging behaviour within the breeding season and between years. A recent study on Arctic Sterna paradise and Common Terns S. hirundo showed that breeding adults can vary both diet and foraging area within a single chick-rearing period (Robertson et al. 2014). I examined this further by comparing the foraging areas used by Black-legged Kittiwakes Rissa tridactyla breeding on Coquet Island (Figure 1), northeast England during two different breeding stages (incubation and chick-rearing) and during the same breeding stage in two consecutive years (chick rearing in 2011 and 2012).
 Figure 1. Aerial view of Coquet Island (55º 20’ N, 1º 32’ W) from the southwest
Figure 1. Aerial view of Coquet Island (55º 20’ N, 1º 32’ W) from the southwest
© Paul Morrison
During incubation in 2012, birds travelled further from the colony than during chick-rearing (Figure 2). Birds tracked during chick-rearing in 2011 and 2012 were also found to forage in different areas in the two years. While birds fed almost exclusively on sandeel during different breeding stages, sandeels consumed during incubation were larger than those eaten during chick-rearing in 2012 (Figure 3). The same environmental variables (such as sea surface temperature, plankton abundance and water depth) did not consistently explain where birds foraged in different breeding stages and years, which suggests that such variables cannot be expected to predict where birds will forage over extended spatiotemporal scales.
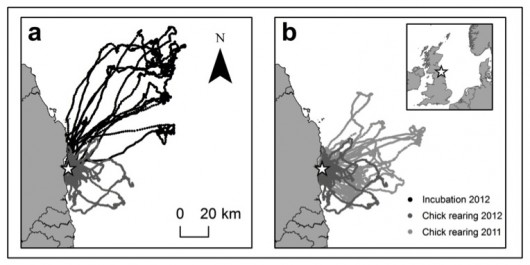 Figure 2. Kittiwake foraging tracks during (a) Incubation and Chick-rearing in 2012 and
Figure 2. Kittiwake foraging tracks during (a) Incubation and Chick-rearing in 2012 and
(b) Chick-rearing in 2011 and 2012 recorded from 23 May to 3 July 2012 and 14 June to 17 June 2011. Coquet Island (55º 20’ N, 1º 32’ W) is represented by a star
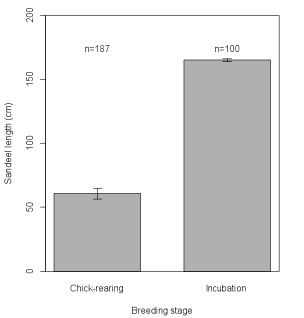
Figure 3. Mean ± standard error length of sandeels (cm) consumed by kittiwakes at different stages of the breeding cycle in 2012
The results of this study show that Kittiwake foraging behaviour and variables associated with foraging locations can change considerably within a single breeding stage and among years. This emphasises the importance of tracking birds over longer time periods when attempting to identify foraging areas for designation as MPAs. Offshore protected areas based on data from a single breeding stage or year are unlikely to represent a large proportion of seabird foraging areas, as areas useful for foraging are highly variable.
Finding ways to reduce the impact of human activities on marine ecosystems are imperative in order to prevent further declines in seabird populations. By tracking seabirds over longer time periods, we can gain a fuller understanding of how birds use the marine environment, and contribute to the conservation of mobile marine predators.
References
Chivers, L.S., Lundy, M.G., Colhoun, K., Newton, S.F., Houghton, J.D.R. & Reid, N. 2013. Identifying optimal feeding habitat and proposed Marine Protected Areas (pMPAs) for the Black-legged Kittiwake (Rissa tridactyla) suggests a need for complementary management approaches. Biological Conservation 164: 73–81. View
Cury, P.M., Boyd, I.L., Bonhommeau, S., Anker-Nilssen, T., Crawford, R.J.M., Furness, R.W., Mills, J.A., Österblom, H., Paleczny, M., Piatt, J.F., Roux, J-P., Shannon, L. & Sydeman, W.J. 2011. Global seabird response to forage fish depletion – one-third for the birds. Science 334: 1703–1706. View
Frederiksen, M., Wanless, S., Harris, M.P., Rothery, P. & Wilson, L.J. 2004. The role of industrial fisheries and oceanographic change in the decline of North Sea Black-legged Kittiwakes. Journal of Applied Ecology 41: 1129–1139. View
Furness, R.W. & Tasker, M.L. 2000. Seabird-fishery interactions: quantifying the sensitivity of seabirds to reductions in sandeel abundance, and identification of key areas for sensitive seabirds in the North Sea. Marine Ecology Progress Series 202: 253–264. View
Kotzerka, J., Garthe, S. & Hatch, S.A. 2010. GPS tracking devices reveal foraging strategies of Black-legged Kittiwakes. Journal of Ornithology 151: 459–467. View
Lewis, S., Wanless, S., Wright, P.J., Harris, M.P., Bull, J. & Elston, D.A. 2001. Diet and breeding performance of Black-legged Kittiwakes Rissa tridactyla at a North Sea colony. Marine Ecology Progress Series 221: 277–284. View
Monaghan, P. 1992. Seabirds and sandeels: the conflict between exploitation and conservation in the northern North Sea. Biodiversity and Conservation 1: 98–111. View
Oro, D. & Furness, R.W. 2002. Influences of food availability and predation on survival of Kittiwakes. Ecology 83: 2516–2528. View
Rindorf, A., Wanless, S. & Harris, M.P. 2000. Effects of changes in sandeel availability on the reproductive output of seabirds. Marine Ecology Progress Series 202: 241–252. View
Robertson, G.S., Bolton, M., Grecian, W.J., Wilson, L.J., Davies, W. & Monaghan, P. 2014. Resource partitioning in three congeneric sympatrically breeding seabirds; foraging areas and prey utilization. The Auk 131: 434–446. View
Stauss, C., Bearhop, S., Bodey, T.W., Garthe, S., Gunn, C., Grecian, W.J., Inger, R., Knight, M.E., Newton, J., Patrick, S.C., Phillips, R.A., Waggitt, J.J. & Votier, S.C. 2012. Sex-specific foraging behaviour in Northern Gannets Morus bassanus: incidence and implications. Marine Ecology Progress Series 457: 151–162. View
Votier, S.C., Bearhop, S., Witt, M.J., Inger, R., Thompson, D. & Newton, J. 2010. Individual responses of seabirds to commercial fisheries revealed using GPS tracking, stable isotopes and vessel monitoring systems. Journal of Applied Ecology 47: 487–497. View
Blog with #theBOUblog
If you want to write about your research in #theBOUblog, then please see here.

