


A need for further research on the influence of large raptors on other predator species and their cascading effects at the ecosystem level
LINKED PAPER
Top-down limitation of mesopredators by avian top predators: a call for research on cascading effects at the community and ecosystem scale. Terraube, J. & Bretagnolle, V. 2018. IBIS. DOI: 10.1111/ibi.12581. VIEW
Predator communities have undergone profound modifications since the 19th century following global human impact
Both top and mesopredators (see Fig. 2 for a description of the different trophic levels) have been heavily persecuted by humans for centuries, altering their abundance and distribution worldwide. Mesopredators may have benefitted at first from the disappearance of large carnivores and top avian predators. However, even though numerous medium-sized raptors and owls have recovered from high persecution levels experienced in the first part of the 20th century, mesocarnivores are still suffering high mortality through hunting and trapping. On the other hand, mesocarnivores have benefitted from global change and the continuous increase in food subsidies from humans. Finally, top predators (both avian and mammalian) species have been recently recolonizing part of deserted areas. These successive modifications have transformed interactions between predator species and among predator and prey communities. However the influence of changes in the structure and composition of predator communities on species occupying lower positions in the food web is still not well understood and requires urgent research.
The role of top predators varies in different ecosystems
There is extensive evidence that top predators are able to control lower-sized predators occupying the same territory. These top-down processes can affect species at lower trophic levels either through direct killing or through indirect behavior changes driven by fear of predation (for example habitats where predation risk is the highest are less used even though they offer abundant food resources, the so-called “fear effect”, Lima 1998). Such top-down processes can even have cascading effects at the ecosystem-level. For example, Gordon et al. (2017) provided evidence that the removal of a top predator, the Dingo Canis lupus dingo, facilitated shrub encroachment in arid Australian ecosystems through a four-level trophic cascade: increased mesopredator abundance (Red Fox Vulpes vulpes and Feral Cat Felis catus) in the absence of Dingoes resulted in suppressed abundance of consumers of shrub seeds and seedlings, rodents and rabbits, respectively. In turn, suppressed abundances of rodents and rabbits in the absence of Dingoes resulted in increased seedling survivorship and decreased seed removal by rodents.
However, most of the evidence comes from areas where top predators reach reasonable densities, mostly in natural habitats, often located within protected areas (Beschta & Ripple 2009).
Several factors constrain the importance of large-scale top-down processes. Among them, human disturbance and higher food abundance linked to agricultural practices in modified ecosystems. Recent research has showed that human presence can affect habitat use and foraging behaviour of top predators and thus dampen the top-down control they exerted on prey species. For example, in Sweden, wolves avoid the most productive forests with high human presence and are not able to control moose browsing intensity in these young plantations (van Beeck Calkoen et al. 2018). There is also increasing evidence that high food abundance can decouple trophic interactions and allow prey to escape predator regulation. For example, forestry may increase forage availability for ungulates by creating clear-cuts and thus weaken the effects of carnivores on their prey. This is also supposedly the case in agricultural areas where vegetation growth is boosted by an intensification of practices, e.g. fertilization.
Finally, most of the research has focused on large carnivores but very few studies have been conducted on top avian predators.
What about large raptors and their functional role?
Top avian predators including large eagles and eagle owls are able to limit other raptor populations through intra-guild predation. For example, the recolonization of the Eurasian Eagle Owl Bubo bubo in Germany has induced increased breeding failure in two sympatric medium-sized raptors, the Northern Goshawk Accipiter gentilis and the Common Buzzard Buteo buteo through direct killing but also through indirect sub-lethal (fear) effects, e.g. abandonment of nestling sites close to Eagle Owl breeding sites (Mueller et al. 2016).
 Figure 1 Eurasian Eagle Owl (left) and Northern Goshawk (right) © Daniel Burgas Riera
Figure 1 Eurasian Eagle Owl (left) and Northern Goshawk (right) © Daniel Burgas Riera
However, the influence of these species on other mesopredators including corvids and mesocarnivores (Red Fox and mustelids) and their cascading effects on primary consumers (small rodents, passerine birds, lagormorphs) is overlooked. Understanding these trophic interactions has important conservation implications.
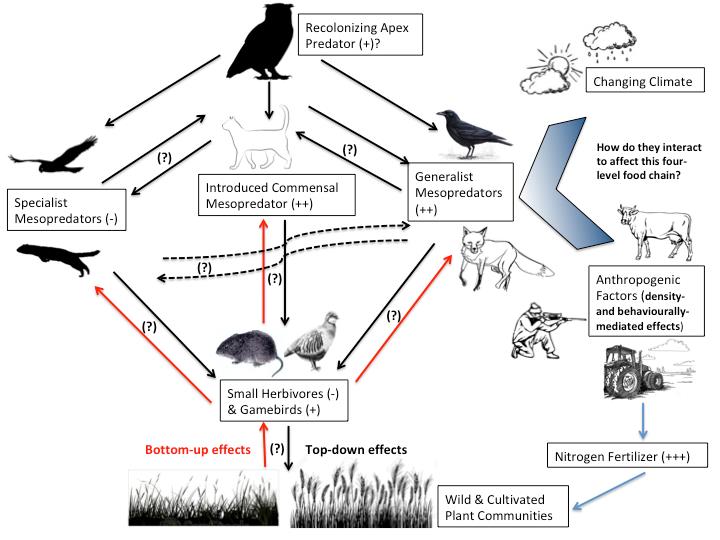 Figure 2 Diagram of hypothesized four-level trophic interactions in European agro-ecosystems highlighting the potential role of recolonizing avian top predators and the factors affecting the structure of this food web (signs in parentheses describe European population trends or levels of resource use)
Figure 2 Diagram of hypothesized four-level trophic interactions in European agro-ecosystems highlighting the potential role of recolonizing avian top predators and the factors affecting the structure of this food web (signs in parentheses describe European population trends or levels of resource use)
Four important future research directions
Both experimental and observational approaches can help deciphering the role of top avian predators in ecosystems. Combining GPS telemetry data and camera trap surveys of large raptors and sympatric mesopredators (mammalian mesopredators and corvids) would allow a better understanding of the behavioral response of these mesopredators to variations in predation risk. Experimental manipulations of fear using visual (raptor decoy) or acoustic cues (call playbacks) of different large raptor species (Billings et al. 2017) would also make it possible to determine whether mesopredators reduce the amount of time spent hunting in areas with cues of large raptor presence and if it alleviates predation pressure from mesopredators on the favored prey species.
Natural recolonization or reintroduction programs of large raptors could be used as natural experiments allowing to better understand how the return of these species influence ecosystems. Golden Eagle Aquila chrysaetos and Eagle Owl are recolonizing part of their former distribution range or are being reintroduced in other areas, offering an opportunity to monitor closely the changes in mesopredator and prey communities before and after the return of these top predators.
There is a need for developing research and science communication on these aspects in tropical ecosystems. Several mesopredator species are considered as pests for their impact on the livelihoods of local communities in numerous countries of the Global South. Emphasizing the potential of large raptors for limiting these mesopredators could be a powerful argument to promote conservation of top avian predators still experiencing high levels of persecution.
 Figure 3 Juvenile Martial Eagle Polemaetus bellicosus in Senegal © Alexandre Villers
Figure 3 Juvenile Martial Eagle Polemaetus bellicosus in Senegal © Alexandre Villers
Towards valuating the regulatory services of top avian predators? Valuating specific ecosystem services provided by animal species is a debated topic. Private hunting estates in several European countries particularly in the U.K. and Spain, bring substantial economic benefits but also experience intensive predator management and wildlife crime. Removing mesopredators is costly and its positive effect on the abundance of game species is controversial, while top avian predators are still suffering illegal persecution in these areas, e.g. Golden Eagles in Scottish grouse moors. Mesopredators are regularly predated by several species of large raptors that could potentially limit mesopredator abundance on hunting estates. However, the effects of top avian predators on these conflicting species (Red Fox and Eurasian Magpie Pica pica for example) and the economic benefits their presence could bring through avoided costs of predator control are still poorly understood.
 Figure 4 Golden Eagle Aquila chrysaetos nest with two fledglings in the French Alps. The prey in the center of the eyrie is a Red Fox © Alexandre Villers
Figure 4 Golden Eagle Aquila chrysaetos nest with two fledglings in the French Alps. The prey in the center of the eyrie is a Red Fox © Alexandre Villers
References
Beschta, R.L. & Ripple, W.J. 2009. Large predators and trophic cascades in terrestrial ecosytems of the western United States. Biol. Conserv. 142: 2401–2414. VIEW
Billings, A.C., Greene, E. & MacArthur-Waltz, D. 2017. Steller’s Jays assess and communicate about predator risk using detection cues and identity. Behav. Ecol. 28: 776–783. VIEW
Gordon, C.E., Eldridge, D.J., Ripple, W.J., Crowther, M.S., Moore, B.D. & Letnic, M. 2017. Shrub encroachment is linked to extirpation of an apex predator. J. Anim. Ecol. 86: 147–157. VIEW
Lima, S.L. 1998. Nonlethal effects in the ecology of predator–prey interactions. Bioscience 48: 25–34. VIEW
Mueller, A.K., Chakarov, N., Heseker, H. & Krüger, O. 2016. Intraguild predation leads to cascading effects on habitat choice, behavior and reproductive performance. J. Anim. Ecol. 85: 774–784. VIEW
van Beeck Calkoen, S.T.S., Kuijper, D.P.J., Sand, H., Singh, N.J., van Wieren, S.E. & Cromsigt, J.P.G.M. 2018. Does wolf presence reduce moose browsing intensity in young forest plantations? Ecography, DOI: 10.1111/ecog.03329. VIEW
Image credit
Featured image: Juvenile Golden Eagle Aquila chrysaetos, Pyrenees © Daniel Burgas Riera
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here.

