

 LINKED PAPER
LINKED PAPER
Spatial and temporal variation in breeding performance in the increasing White-tailed Eagle Haliaeetus albicilla population to the east of the Baltic Sea. Dementavičius, D., Rumbutis, S. & Treinys, R. 2022 Bird Study. doi: 10.1080/00063657.2022.2124954 VIEW
The White-tailed Eagle (Haliaeetus albicilla) is the largest diurnal raptor in Europe. Its populations have been recovering over the past few decades in different areas of it distribution range following environmental improvements to reduce the contamination by DDT pesticides after their ban and decreased persecution (Helander & Stjernberg 2003).
What should we expect during population increases?
New pairs establish their territories near conspecifics in the same habitats, also in different (Ekblad et al. 2020) or even in less suitable habitats (Heuck et al. 2017). This process results in habitat heterogeneity and increased prey abundance which reduces the size of individual nesting territories. The territorial behaviour of White-tailed Eagle toward conspecifics peaks during the nestling hatching and rearing periods between April and up until May (Krone et al. 2013). Moreover, intraspecific fighting may end in the death of adult birds (Saurola et al. 2003). Therefore, we expected that increases in the number of pairs may lead to negative effects on breeding performance due to limited access to and/ or poorer resources or less time available to spend on nestling provisions because of more intense territorial disputes.
Material for the study
Data were collected in two study areas, coastal and inland, located in the eastern region of the Baltic Sea, Lithuania, Central Europe. Field work was done by my colleagues Dr. Deivis Dementavičius and Saulius Rumbutis, former being the leading researcher of the White-tailed Eagle in Lithuania since the end of 1990s. For this study, we analysed data collected between 2005 and 2020 on the number of pairs, their nesting success and nestling measurements, which were taken during ringing.

Figure 1 Successful nests were climbed once a season © Andrius Keraitis.
Increases in the number of pairs
For 16 years the size of the local population in the coastal area had increased from 12 to 42 pairs, and in the inland area from six to 23 pairs. The mean nearest distance between the nests of neighbouring pairs in the coastal area in 2005 was ca. 7 km, which by 2020 had decreased to ca. 2 km. These territories are located very close together considering the large home-ranges that are characteristic of White-tailed Eagles. In the inland area, pairs were spaced at an average of ca. 20 km in 2005, decreasing to ca. 9 km in 2020. In conclusion, a three-fold increase in the local population numbers resulted in ca. a three-fold compression of territories per pair in both study areas.
Overview of reproduction parameters
Between 2005 and 2020, White-tailed Eagles raised at least one fledging in 381 (62.8%) out of an observed 606 occupied nests. The average number of nestlings was 1.07 per occupied nest (n = 606), and the average number of nestlings per successful brood was 1.70 (n = 381). Out of 448 nestlings, 48.7% were sexed as females, based on tarsus diameter (following the morphological method described in Helander et al. 2007). These figures are similar to White-tailed Eagle reproduction parameters reported in other regions around the Baltic Sea and may be regarded as being close to the natural reproduction level, estimated before 1950 (Helander et al. 2013). The ages of the nestlings during ringing were, on average, 48 days (n = 448), with individuals sexed as males weighing, on average, 3.79 kg (n = 230) and females 4.61 kg (n = 218).

Figure 2 The number of nestlings in the brood was estimated, nestlings were then weighed, wing length and tarsus thickness measured prior to ringing © Rimgaudas Treinys.
Breeding performance fluctuated but did not decrease
All the reproductive variables analysed (i.e., nesting success, brood size, proportion of females among nestlings, nestling body condition index) fluctuated strongly annually. However, no statistically significant decrease was found as we expected. We also tested the variation of the mean annual number of nestlings per pair as well as its coefficient of variation separately for the coastal and inland study areas, where we also found no significant changes over time.
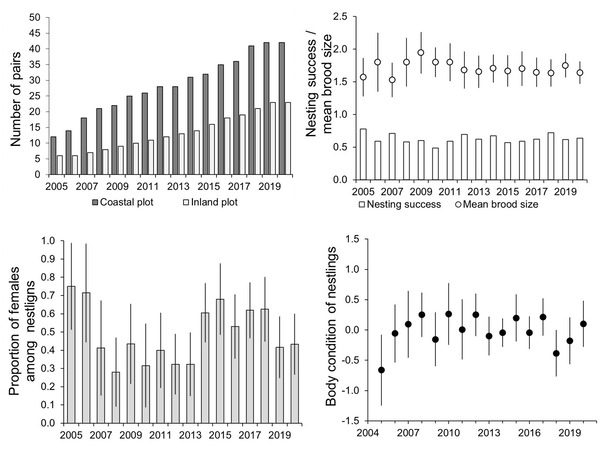
Figure 3 Annual variation in White-tailed Eagle pair numbers, nesting success (i.e., proportion of successful pairs) (bars) and mean brood size (circles), proportion of females among nestlings, mean body condition of nestlings in the eastern region of the Baltic Sea. Error bars represent 95% confidence intervals.
Weak habitat effect on breeding performance
In the coastal area, nesting territories were assigned either to a ‘lagoon’ habitat, or to a ‘delta’ habitat. In the inland study area, the nesting territories were assigned either located in an ‘inland fishpond’ habitat, or located in an ‘inland natural’ habitat. The probability of successful nesting, nestling sex and number of nestlings in a brood were found to be similar across pairs, associated with the aforementioned habitats, representing the wide-ranging environmental gradient in the region. Only nestling body condition was found to be habitat-dependent: the best body condition achieved in nestlings reared in the ‘lagoon’ habitat in a coastal study area.

Figure 4 View from the nest of a White-tailed Eagle pair. Most reproduction parameters were similar across different habitats © Rimgaudas Treinys.
Conclusions
In summary, we did not find any support for deterioration in measured reproduction parameters in the population, despite ongoing increases in the local environment, characterised by different types of feeding habitats. Of note, our finding is applicable only for the population level and does not automatically negate effect of conspecific density at individual nesting territory level and this topic is the aim of our next study.
The most plausible reason is that availability of food resources has not been depleted during periods of population increase. This is supported by the results of several our recent studies. First, there have been no signs that, during a population increase, eagles started to exploit less profitable, alternative prey that may indicate food limitation (Dementavičius et al. 2020). Second, the predation on Northern Pike (Esox lucius) – a keystone prey species in the Baltic Sea region – increased significantly between 2005 and 2018, most likely as a functional response of eagles to the increased pike availability because of the large-scale pike stocking programme in Lithuanian freshwaters and improved control of recreational fishery (Virbickas et al. 2021).
Future research
Retained reproduction rates will most likely result in further population growth in the region located east of the Baltic Sea, at least in short- and mid-term perspectives. The larger population consisting of territorial adults and vagrant immature birds may potentially lead to more frequent interspecific interactions. Therefore, research on this species’ feeding behaviour and possible non-lethal effects, e.g., nesting habitat displacement, supressed feeding activities of coexisting prey and non-prey species (Sergio & Hiraldo 2008), will be important to understand the functions and consequences of a recovered avian predator in local ecosystems.
References
Dementavičius, D., Rumbutis, S., Virbickas, T., Vaitkuvienė, D., Dagys, M. & Treinys, R. 2020. Spatial and temporal variations in the White-tailed Eagle Haliaeetus albicilla breeding diet revealed by prey remains. Bird Study 67: 206-216. VIEW
Ekblad, C., Tikkanen, H., Sulkava, S. & Laaksonen, T. 2020. Diet and breeding habitat preferences of White-tailed Eagles in a northern inland environment. Polar Biology 43: 2071-2084. VIEW
Helander, B., Bignert, A., Herrmann, C. & Stjernberg, T. 2013. White-tailed Eagle productivity. HELCOM Core Indicator Report. VIEW
Helander, B., Hailer, F. & Vilà, C. 2007. Morphological and genetic sex identification of white-tailed eagle Haliaeetus albicilla nestlings. Journal of Ornithology 148: 435-442. VIEW
Helander, B. & Stjernberg, T. 2003. Action plan for the conservation of White-tailed Sea Eagle (Haliaeetus albicilla). BirdLife International report to the Bern Convention on the Conservation of European Wildlife and Natural Habitats, Strasbourg.
Heuck, C., Herrmann, C., Schabo, D.G., Brandl, R. & Albrecht, J. 2017. Density-dependent effects on reproductive performance in a recovering population of White-tailed Eagles Haliaeetus albicilla. Ibis 159: 297-310. VIEW
Krone, O., Nadjafzadeh, M. & Berger, A. 2013. White-tailed Sea Eagles (Haliaeetus albicilla) defend small home ranges in north-east Germany throughout the year. Journal of Ornithology 154: 827-835. VIEW
Saurola, P., Stjenberg, T., Högmander, J., Koivusaari, J., Ekblom, H. & Helander, B. 2003. Survival of juvenile and sub-adult Finnish White-tailed Sea Eagles in 1991–1999: A preliminary analysis based on resightings of colour-ringed individuals. In Helander, B., Marquiss, M. & Bowerman, B. (eds.) Sea Eagle 2000: 155-167. Swedish Society for Nature Conservation, Stockholm.
Sergio, F. & Hiraldo, F. 2008. Intraguild predation in raptor assemblages: a review. Ibis 150: 132-145. VIEW
Virbickas, T., Dementavičius, D., Rumbutis, S., Vaitkuvienė, D., Dagys M. & Treinys, R. 2021. Understanding recreational targets and ecological consequences: increased northern pike stocking reflected in top avian predator diet. European Journal of Wildlife Research 67: 5. VIEW
Image credit
Top right: White-tailed Eagle (Haliaeetus albicilla) nestling © Rimgaudas Treinys.
If you want to write about your research in #theBOUblog, then please see here.

