


LINKED PAPER
Vocal behaviour, parasitic infection, chronic stress and body condition in rufous-naped wrens (Campylorhynchus rufinucha). Meza-Montes, E., Fernández-Gómez, R. A., Llanes-Quevedo, A., Navarro-Sigüenza, A. G., Santiago-Alarcon, D., & Sosa-López, J. R. 2023. IBIS. DOI: 10.1111/ibi.13130. VIEW
The concept of sexual selection was first proposed by Charles Darwin over 150 years ago, and it has since been found to take many forms across different taxa. A relatively more recent concept is that of parasite-mediated sexual selection, in which mate choice is influenced by the effects of parasites upon secondary sexual characteristics such as ornaments or displays, e.g. acoustic signalling. So far, few studies have investigated associations between vocal behaviour and parasitic infections in birds, and these have generally shown inconsistent results.
In a recent study in Ibis, Estefanía Meza-Montes and colleagues examined haemosporidian infection and vocal behaviour, among other traits, in Rufous-naped Wrens (Campylorhynchus rufinucha), to expand our understanding of these interactions.
Parasite-mediated sexual selection hypotheses
The association between the expression of secondary sexual characteristics, such as bird song, and parasitic infection has been suggested to be explained by four potential hypotheses. The first is the Hamilton and Zuk (resistance gene) hypothesis, which suggests that where hosts have coevolved with parasites females prefer males with more elaborate displays as these indicate the presence of resistance genes which can be passed on to offspring (Hamilton & Zuk 1982). The second is the contagion indicator hypothesis, which suggests that females prefer males with elaborate displays because this indicates lower parasite infestation in the males and hence lower risk of acquiring the parasites themselves (Able 1996). The third is the efficient parenting hypothesis, which suggests that females choose males with elaborate displays as being able to maintain these indicates that the males might provide greater parental care (Hamilton 1990). Finally, the parasite avoidance hypothesis suggests that females choose parasite-free males to reduce the probability of contracting the parasites themselves (Borgia & Collis 1989).
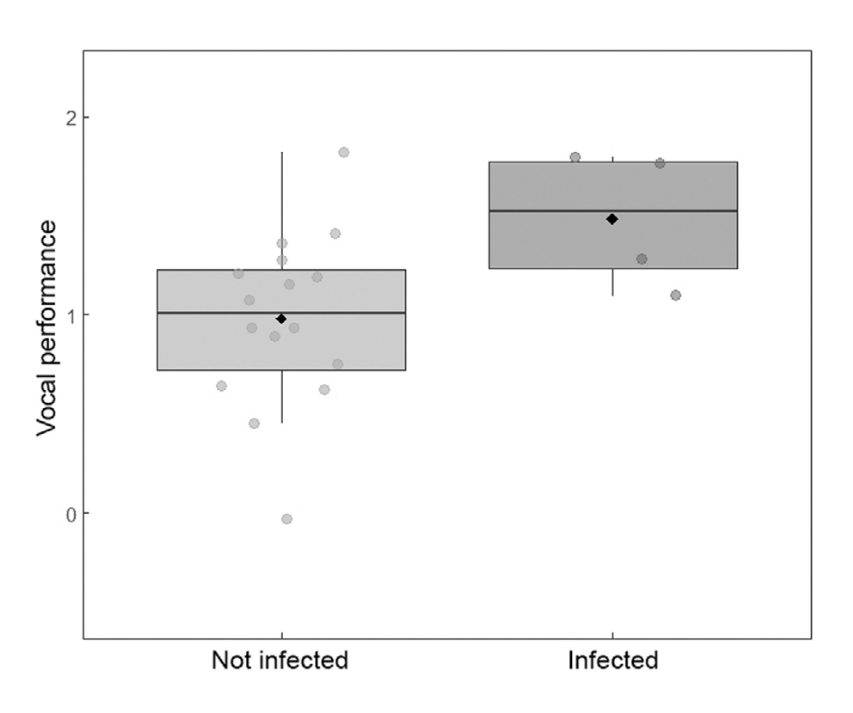
Figure 1. Box plot showing how vocal performance (as measured by number of song bouts per minute) differs according to infection status. Box plots show the median, first (lower) and third (upper) quartile, and whiskers denote values that are not outliers. Dots show the raw data and the black diamond symbols indicate the mean.
Infection, body condition, and songs
The researchers captured 50 Rufous-naped Wrens from 32 territories in Mexico and analysed their infection status, parasite load, body condition, heterophil:lymphocyte (H:L) ratio (index of chronic stress), song complexity (repertoire size), song structural characteristics, and vocal performance (singing rate). The results showed no evidence of an association between infection and body condition in this species, while other studies have shown contrasting results. This could be due to numerous factors such as study differences, the multiple complex factors affecting body condition, or a bias where the infected birds observed in the study were only those that survived due to be being of high quality. There was also no evidence of an association between parasitism and H:L ratio, which may be because haemosporidian parasites had a small or condition-dependent effect.
A key finding was the lack of an association between repertoire size and parasite infection, which doesn’t support suggestions that infection may affect the brain development necessary for the expression of bird songs (Nottebohm 2005). Results also showed that infected birds produced higher song rates than healthy birds, which could be due to biological traits, coevolution with local parasites, or temporal variation during the study period. Finally, infection status and parasite load were found to be correlated between members of a pair, supporting both the contagion indicator and parasite avoidance hypotheses, although the positive relationship between infection status and song rate makes it difficult to understand how vocal behaviours could be being used by females as a cue to avoid infection.
Due to the paucity of research into parasitic infection and acoustic signalling associations in birds, as well as contrasting results across the few existing studies, the relationships and mechanisms remain unclear. The researchers suggest that further work involving larger sample sizes, experimental infections, and additional variables such as age and survival rate could help to improve our understanding of the relationships between parasitic infection, bird health, and vocal behaviour.
References
Able, D.J. (1996). The contagion indicator hypothesis for parasite-mediated sexual selection. Proceedings of the National Academy of Sciences of the United States of America 93: 2229-2233. VIEW
Borgia, G. & Collis, K. (1989). Female choice for parasite-free male satin bowerbirds and the evolution of bright male plumage. Behavioral Ecology and Sociobiology 25: 445-454. VIEW
Hamilton, W.D. (1990). Mate choice near or far. American Zoologist 30: 341-352. VIEW
Hamilton, W.D. & Zuk, M. (1982). Heritable true fitness and bright birds: a role for parasites? Science 218: 384–387. VIEW
Nottebohm, F. (2005). The neural basis of birdsong. PLoS Biology 3: e164. VIEW
Image credits
Top right: Rufous-naped Wren (Campylorhynchus rufinucha) | Charlie Jackson | CC BY 2.0 Wikimedia Commons
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here

