

 LINKED PAPER
LINKED PAPER
Assessing the reproductive consequences of mate retention and pair bond duration in Thorn-tailed Rayadito (Aphrastura spinicauda), a short-lived, socially monogamous Neotropical bird. Botero-Delgadillo, E., Vásquez, R.A. & Kempenaers, B. 2023 Ibis. doi: 10.1111/ibi.13183 VIEW
In socially monogamous birds, long-term pair bonds are known to provide some benefits to individuals that are able to remain together for successive breeding seasons. Retaining a mate can reduce the time and energy required to find a new partner but also increase familiarity and coordination in shared activities. Most evidence supporting the apparent existence of such benefits comes from studies on long-lived bird species, which can remain together for decades. However, what about short-lived, socially monogamous birds, such as most songbirds? According to scientific literature, evidence for the benefits of mate retention in songbirds is controversial. Some studies have reported that remated pairs breed earlier in Eurasian Blue Tits (Cyanistes caeruleus) or Great Tits (Parus major) (Pampus et al. 2005, Culina et al. 2020), whereas others did not find any apparent benefits (e.g., Llambías 2008). Furthermore, some studies have suggested that early breeding or any other measure suggesting increased breeding productivity as a consequence of remating in songbirds may be a by-product of individual breeding experience and age.
In general, most songbirds not only have shorter lifespans than, for example, Macaws or Cranes, but also tend to have lower survival rates. If life is short and survival is low, why would there be a reproductive benefit of remating if mate retention is a rare event in songbirds? Well, not quite. Songbirds in the Neotropics tend to exhibit higher survival rates than their counterparts at temperate latitudes (Scholer 2020). Furthermore, extensive bi-parental care and relatively long-term pair-bonds seem to be typical in some bird families endemic to the Neotropics, such as the Thamnophilidae (antbirds) and Furnariidae (ovenbirds and allies). Therefore, we thought that if mate retention could confer some benefits to songbirds, a species belonging to any of these groups would be a great candidate to assess this. We therefore decided to study the Thorn-tailed Rayadito (Aphrastura spinicauda), a 11 g socially monogamous furnariid that breeds during the Austral spring (September–December) in tree cavities in the humid forests across Chile and in western Argentina. Rayaditos are an excellent study system since they use nestboxes whenever offered.
We studied an isolated population of Thorn-tailed Rayadito in Bosque Fray Jorge National Park (30°38’S, 71°40’W), located in the Coquimbo Region, north-central Chile. This region is mainly covered by a semi-arid steppe, but fragmented relicts of Valdivian temperate forest persist atop the coastal mountain range (~500–600 m) due to an oceanic fog-induced microclimate that maintains humid conditions throughout the year [Figure 1].

Figure 1 Panoramic view of Bosque Fray Jorge National Park, north-central Chile, with dominant semi-arid steppe. The oceanic fog atop the coastal mountain range where forest fragments are located can be observed at the background © Esteban Botero-Delgadillo.
In Fray Jorge, Rayaditos only breed inside forest fragments that are surrounded by a shrub-dominated matrix, and lay only one clutch of 1–4 eggs during a breeding season. We used data from a long-term breeding monitoring program that started in 2007 and that has served to annually collect data on the reproductive biology of Rayaditos. For this study in particular, we used a 10-year dataset (2008–2017) that contained information for 159 unique breeding pairs and 243 breeding attempts (there were more breeding attempts because one pair could breed for more than one year). Data were collected by monitoring 101–157 nestboxes that were regularly distributed within forest fragments [Figure 2].
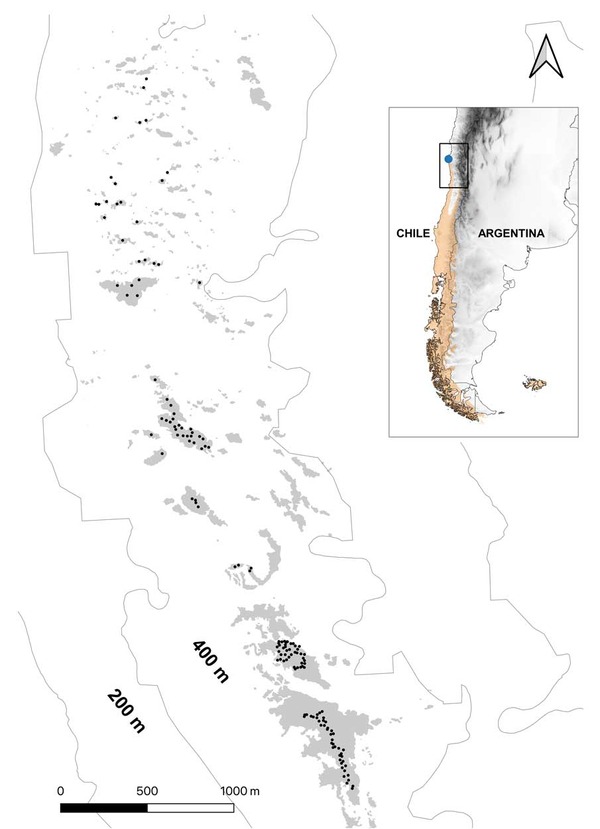
Figure 2 Distribution of nestboxes installed since 2007 in Bosque Fray Jorge National Park, north-central Chile, to study the breeding biology of Thorn-tailed Rayadito (Aphrastura spinicauda). Natural fragments of forest relicts atop the coastal mountain range are shown in light grey. The small panel on the right shows the species’ distribution and the location of the study area.
What were our goals and what did we find?
We first wanted to describe the mating patterns in the study population, including the duration of pair bonds, the percentage of newly formed versus remated pairs, and the percentage of mate loss versus divorce (defined as cases where both former breeding pair members are known to have survived) as a cause of pair dissolution. We found that nearly 30% of all breeding pairs bred together for at least two consecutive years, and some mated for six years [Figure 3]. We also found that 40–60% of all breeding attempts were made by remated pairs. Among all birds that changed their mate between years, mate loss was the prevailing cause of pair dissolution over divorce (27 females: 30% divorcees and 70% widows; 40 males: 22% divorcees and 78% widows).
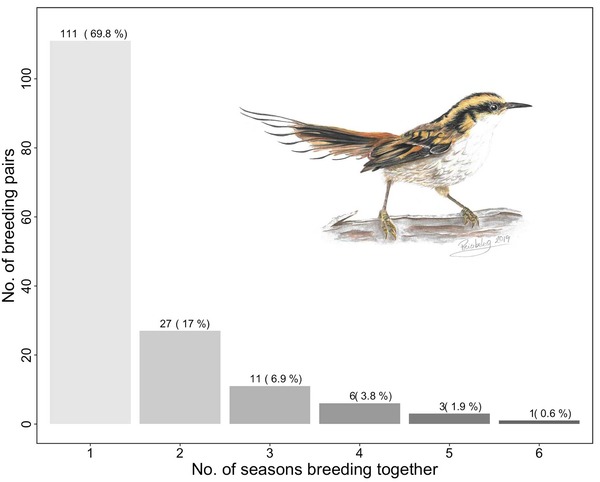
Figure 3 Frequency distribution of the number of seasons breeding together for 159 breeding pairs of Thorn-tailed Rayadito during 2009–2017 in Bosque Fray Jorge National Park, north-central Chile. Illustration of Thorn-tailed Rayadito © Priscila Escobar Gimpel.
Our study also aimed at determining whether reproductive success was higher for remated pairs than for newly-formed pairs, whether mate replacement did cause a subsequent reduction in reproductive success for breeding individuals, and whether successive remating, namely the same pair breeding in two or more consecutive years, influenced reproductive success. In short, our analyses suggested that newly formed pairs laid eggs later (2–10 days) and had slightly smaller clutches (0.1–0.5 eggs/breeding season) than remated pairs. Also, we found that females laid on average 0.2–0.7 fewer eggs after changing their previous partner due to mate loss. Moreover, females from remated pairs seemingly laid eggs earlier and laid larger clutches from the second breeding attempt onwards [Figure 4].
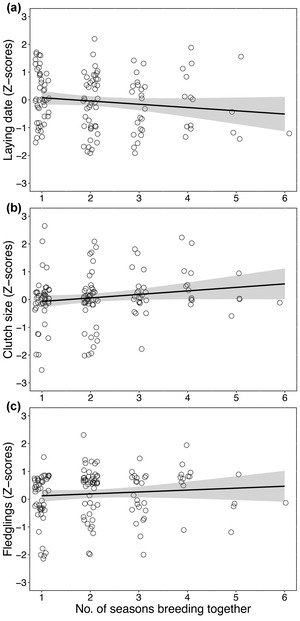
Figure 4 Relationship between the number of seasons a pair bred together and measures of reproductive success in Thorn-tailed Rayadito during 2009–2017 in Fray Jorge National Park, north-central Chile. Panels show raw data for (a) laying date, (b) clutch size, and (c) number of fledglings produced. Model predictions are depicted with 95% confidence intervals (lines and shaded area).
Our results suggest that in Rayaditos, newly-formed pairs seem less efficient in reproduction. Furthermore, it seems that minor yearly reproductive benefits of mate retention might accumulate for individuals that are able to breed with the same partner over many years. These apparent advantages of extended pair bonds were still supported by our data after controlling for the potential effects of individual age and breeding experience in our analyses.
Because breeding habitat is limited in our study population, Rayaditos could benefit from remating if the number of individuals that can breed exceeds the number of available breeding positions. This is likely the case, as Fray Jorge is a densely populated environment where adult apparent survival is close to 70%, remating is frequent, and individuals may be forced to delay their first breeding attempt (Botero-Delgadillo et al. 2017, 2019). We think that low adult mortality possibly increases the probability of retaining a mate over several breeding seasons, and that competition for nesting sites and breeding partners may promote breeding with the same partner across years. If high survival and competition for breeding positions is frequent across the tropics, profitable long-term pair bonds might be more common in tropical songbirds than previously considered.
References
Botero-Delgadillo, E., Quirici, V., Poblete, Y., Cuevas, E., Kuhn, S., Girg, A., Teltscher, K., Poulin, E., Kempenaers, B. & Vásquez, R.A. 2017. Variation in fine-scale genetic structure and local dispersal patterns between peripheral populations of a South American passerine bird. Ecology and Evolution 7: 8363-8378. VIEW
Botero-Delgadillo, E., Quirici, V., Poblete, Y., Cuevas, E., Kuhn, S., Girg, A., Poulin, E., Kempenaers, B., & Vásquez, R.A. 2019. Ecological and social correlates of natal dispersal in female and male Thorn-tailed Rayadito (Aphrastura spinicauda) in a naturally isolated and fragmented habitat. Auk 136: ukz016. VIEW
Culina, A., Firth, J.A. & Hinde, C.A. 2020. Familiarity breeds success: pairs that meet earlier experience increased breeding performance in a wild bird population. Proceedings of the Royal Society B 287: 20201554. VIEW
Llambías, P.E., Wrege, P. & Winkler, D.W. 2008. Effects of site fidelity and breeding performance on mate retention in a short-lived passerine, the tree swallow Thachycineta bicolor. Journal of Avian Biology 39: 493-499. VIEW
Pampus, M., Schmidt, K.-H. & Wiltschko, W. 2005. Pair bond and breeding success in Blue Tits Parus caeruleus and Great Tits Parus major. Ibis 147: 92-108. VIEW
Scholer, M.N, Strimas-Mackey, M. & Jankowski, J.E. 2020. A meta-analysis of global avian survival across species and latitude. Ecology Letters 23: 1537-1549. VIEW
Image credit
Top right: Thorn-tailed rayadito (Aphrastura spinicauda) © Silvia Lazzarino.
If you want to write about your research in #theBOUblog, then please see here.

