

 LINKED PAPER
LINKED PAPER
When birds sing at the same pitch, they avoid singing at the same time. Chronister, L.M., Rhinehart, T.A. & Kitzes, J. 2023 Ibis. doi: 10.1111/ibi.13192 VIEW
For animals that communicate with each other using sound, one challenge has long been recognized: in noisy conditions, an individual’s acoustic signals may not be heard due to interference by other sounds. Readers will be familiar with this problem if they have ever tried to talk to another person in a crowded room and in fact, this phenomenon has been aptly named the “cocktail party problem” (Bee & Micheyl 2008). Readers may also remember certain individuals whose voices seem to carry further than others in these conditions, and ways they have themselves tried to counter the cocktail party problem—perhaps by raising their voice, speaking in a slower, clearer manner, or by focusing all attention on just their conversational partner’s voice. Similarly, strategies for dealing with the cocktail party problem have been recognised in many species of animals such as frogs (Duellman & Pyles 1983), insects (Sueur 2008), and birds (Hart et al. 2021).
Birds face many sources of potential interference when they try to communicate using sound. Not only can members of their own species contribute to the problem, but also other sound producing animals, anthropogenic (human-caused) noise, and natural environmental noise such as water or wind can be significant sources of interference. Further still, many songbirds take part in dawn choruses—a time period right around sunrise in which numerous different bird species in a community all try to sing much more than during other parts of the day. The particular source of sound interference will define the way that birds respond to the problem of acoustic interference.
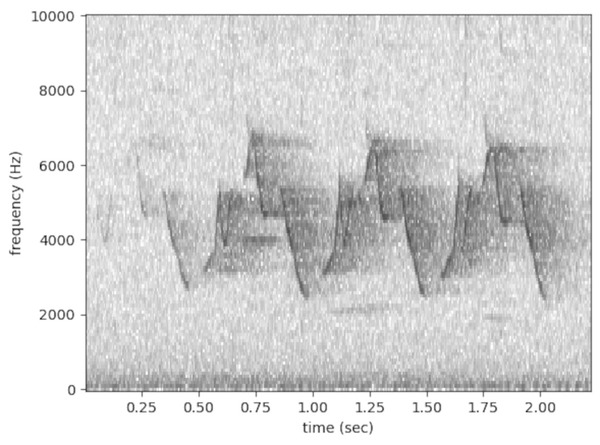
Figure 1 An example of a Common Yellowthroat (Geothlypis trichas) song displayed as a spectrogram. Common Yellowthroat is one of the species studied in Chronister et al. (2023). Spectrograms are graphical representations of sound with time on the X axis, frequency (pitch) on the Y axis, and darker regions representing louder sounds. The audio used to produce this spectrogram comes from Chronister et al. (2021), the same publication which supplied the data used in Chronister et al. (2023). The spectrogram was produced using Opensoundscape (http://opensoundscape.org/en/latest/).
How birds avoid interference
One of the simplest strategies birds use to deal with the cocktail party problem when they’re faced with numerous species or individuals all wanting to make sound at once is to wait for their turn. Birds have been observed to pause their singing when the buzz of cicadas starts up (Hart et al. 2015), cycle longer singing bouts with other species (Cody & Brown 1969), or even wait until brief pauses between the individual songs of other birds to give their own song (Brumm 2006). Singing can be a costly activity for birds both in terms of the energy expenditure of singing itself and in terms of time they could be spending doing other important things, but it’s also a vital activity for territorial defense and mate attraction. Thus, for birds, waiting until they have an opportunity to be clearly heard can pay off.
Decisions about when to sing are not the only means by which birds tackle the cocktail party problem. Other aspects of each species’ songs can enable them to sing more while worrying less about timing their songs just right. For instance, different bird species tend to sing at different pitches, a characteristic that is influenced by body and bill size (Demery et al. 2021). Two songs occurring at the same time but at different pitches (i.e., frequencies) will not necessarily interfere with each singer being heard. Thus, birds may only need to pay attention to timing their songs with other birds that share their pitch. This strategy has been demonstrated previously where birds were shown to segregate their singing into separate time periods more often when they sang in the same frequency “channels” (Planqué & Slabbekoorn 2008).

Figure 2 An example of an audio clip containing the songs of Common Yellowthroat and several other bird species demonstrating the challenge of singing in crowded conditions. The audio used to produce this spectrogram comes from Chronister et al. (2021), the same publication which supplied the data used in Chronister et al. (2023). The spectrogram was produced using Opensoundscape (http://opensoundscape.org/en/latest/).
Decisions on timing according to pitch
Frequency channels are not clear-cut, however, and in some communities such as the one we studied, there can be many instances of intermediate amounts of overlap in pitch between the songs of different bird species. Further still, the range of pitch different species’ songs occupy also vary greatly such that one species in any pair might be entirely overlapped whereas the other is only overlapped in a small portion of its range. So how does a bird make a decision about when to avoid singing over another bird when overlap in pitch is not necessarily binary? We hypothesised that the decision to overlap or not overlap would also not be binary such that birds would instead be increasingly more likely to avoid song overlap in time as their pitches increasingly overlapped.
We put our hypothesis to the test using recordings from a community of birds living in the temperate forests of Pennsylvania, USA (Chronister et al. 2021). We used computer simulations of what would happen if birds were singing randomly in time with respect to other bird species to compare the outcome with that of our actual data. Ultimately, we found support for our hypothesis: birds with more overlap in pitch typically sang over one another far less than in simulations whereas this was not the case for species with relatively little overlap in pitch. The pattern was that of a linear correlation. This suggests that birds in this community are paying closer attention to avoid singing over other birds that have the greatest risk of interfering with their songs in terms of pitch, and will still avoid singing over other species with relatively less overlap in pitch but to a lesser degree.
References
Bee, M. A., & Micheyl, C. 2008. The cocktail party problem: What is it? How can it be solved? And why should animal behaviorists study it? Journal of Comparative Psychology 235-251. VIEW
Brumm, H. 2006. Signalling through acoustic windows: nightingales avoid interspecific competition by short-term adjustment of song timing. Journal of Comparative Physiology A 1279-1285. VIEW
Chronister, L.M., Rhinehart, T.A., Place, A. & Kitzes, J. 2021. An annotated set of audio recordings of Eastern North American birds containing frequency, time, and species information. Ecology e03329. VIEW
Cody, M.L. & Brown, J.H. 1969. Song Asynchrony in Neighboring Bird Species. Nature 778-780. VIEW
Demery, A.C., Burns, K.J. & Mason, N.A. 2021. Bill size, bill shape, and body size constrain bird song evolution on a macroevolutionary scale. Ornithology ukab011. VIEW
Duellman, W.E. & Pyles, R.A. 1983. Acoustic Resource Partitioning in Anuran Communities. Copeia 639-649. VIEW
Hart, P.J., Ibanez, T., Paxton, K., Tredinnick, G., Sebastián-González, E. & Tanimoto-Johnson, A. 2021. Timing is Everything: Acoustic Niche Partitioning in Two Tropical Wet Forest Bird Communities. Frontiers in Ecology and Evolution 753363. VIEW
Hart, P.J., Hall, R., Ray, W., Beck, A. & Zook, J. 2015. Cicadas impact bird communication in a noisy tropical rainforest. Behavorial Ecology 839-842. VIEW
Planqué, R. & Slabbekoorn, H. 2008. Spectral Overlap in Songs and Temporal Avoidance in a Peruvian Bird Assemblage. Ethology 262-271. VIEW
Sueur, J. 2008. Cicada acoustic communication: potential sound partitioning in a multispecies community from Mexico (Hemiptera: Cicadomorpha: Cicadidae). Biological Journal of the Linnean Society 379-394. VIEW
Image credit
Top right: Common Yellowthroat (Geothlypis trichas) © Patrice Bouchard.
If you want to write about your research in #theBOUblog, then please see here.

