

Linked paper
Reducing bycatch in gillnets: A sensory ecology perspective.
Graham R. Martin & Rory Crawford. 2014. Global Ecology and Conservation. DOI: 10.1016/j.gecco.2014.11.004
In 2013, the first global review of seabird bycatch in gillnet fisheries was published. It estimated, conservatively, that 400,000 birds were killed every year in this gear (Zydelis et al, 2013), making it responsible for more mortalities than longline and trawl fisheries combined (estimated to kill 300,000 birds/year) (Anderson et al, 2011).
Reducing this toll should be a conservation priority for several reasons. The clear moral imperative aside, minimising bycatch of non-target species is a fundamental aspect of sustainable fishing, as recognised by the FAO (FAO, 1995).Further, seabirds are the most threatened group of birds globally, and bycatch, as one of the major threats to seabirds (as well as a broadly unwelcomed by-product of fishing), is an obvious target for conservation efforts (Croxall et al, 2012). Finally, it is already known that the magnitude of bycatch in some locations is sufficient to result in population declines, for example at least two Baltic Sea seabird populations (Greater Scaup and Common Guillemot) are thought to be declining because of bycatch (Zydelis et al, 2009).
So the case for doing something is fairly clear-cut – but what should we be doing? Over the past two years, I have discussed ideas about the reduction of seabird bycatch in gillnets with lots of conservationists, and all sorts of things– different coloured twines, lights, different kinds of floats on nets – were mooted. Although many of these ideas may have merit, I, and others, were probably falling into the classic trap of looking at the problem from a human perspective. Really, we had to look at this from a bird’s eye view, and this is what led me to get in touch with Graham Martin about what birds can actually see underwater.
Graham undertook a review (Reducing bycatch in gillnets: A sensory ecology perspective), of what is known of bird – and other bycatch-prone taxa – sensory capacities. As vision is the primary source of information gathering for most vertebrates, this was the main focus of the review (though echolocating cetaceans are a notable and bycatch-prone exception). With a conservation (rather than a sensory ecology) audience in mind, the review goes back to the fundamentals of vision – how light behaves in water, the physical constraints on underwater sight, and the trade-offs involved for animals with eyes that are capable of seeing in terrestrial and aquatic contexts.
So, how well do birds see underwater?
While, as already noted, it is not particularly helpful to view this through a human perspective, we must also not assume superhero levels of sight in birds foraging underwater. This is a tricky visual environment, posing many challenges for vision.
The first challenge for seeing underwater is the amount of light penetrating below the surface. Light is reflected (and thus ‘lost’) from the water’s surface. Light that does enter the water column is quickly scattered and absorbed; some wavelengths are absorbed before others, meaning a very narrow band of wavelengths are available for vision at depth. This means that many diving seabirds capable of diving deeper than 100m (e.g. common guillemots, king penguins), forage during the day at depths that mean they are effectively ‘nocturnal’ foragers, feeding in conditions equivalent to twilight or moonlight.
Next, the challenge of seeing in both air and water. You will likely have witnessed this first hand when going for a swim and taking a look around underwater – your vision is blurry. This occurs because, in eyes that are adapted for the terrestrial environment, the cornea at the front of the eye provides a curved interface between the air and the fluid-filled chamber inside the eye. This curvature focuses whatever you are looking at onto the retina – but when the medium changes from air to water this refractive power of the cornea is lost because the water outside the eye is similar in nature to the fluid inside the eye. The more powerful a cornea is – i.e. the more curved it is – the stronger the blurring effect when water is entered.
There is an even starker trade-off between the sensitivity – the ability of any eye to ‘collect’ light – and spatial resolution – the ability to detect the ‘details’ of a given scene. Thus an eye that is highly sensitive (i.e. it can see at low light levels), is fundamentally unable to detect fine details in brighter conditions. This clearly has consequences for diving birds – the fine nylon mesh of gillnets clearly falls into the ‘fine details’ category. But how much detail can birds actually see? The acuity – a simple measure of the ability to see detail – has only been directly measured for one bird species underwater: Great Cormorants (White et al, 2007). Acuity can be measured by determining an animal’s ability to discern different levels of contrast. These experiments indicated that cormorant vision is actually rather poor underwater – the contrast of prey fish against the background has a major impact on a cormorant’s ability to detect it (see Fig 1). The belief that diving seabirds locate prey from a distance and then hunt it down is thus likely a fallacy – at very least for cormorants. They are instead reliant on random searching followed by prey capture at close quarters (White et al, 2007).

Fig. 1. Prey detectability model for a Great Cormorant based upon the data of Figures 2 and 3 demonstrating the effects of contrast and viewing distance. The model is based upon a Great Cormorant foraging on a capelin Mallotus villosus (10 cm total length) type of fish at an ambient illumination of 10 lux, which has a contrast of 90, 60 and 30% viewed from a distance of 0.63, 1.05 or 2.12 m. From White et al. (2007).
All of this would seem to make the search for bycatch mitigation measures in gillnets extremely difficult – how can we reduce the bycatch of animals that don’t actually see particularly well underwater – and still ensure that fishermen can catch fish? Faced with this conundrum, minds often turn to the fact that some birds (some gulls, passerines, parrots and ostriches) can see into the UV part of the visual spectrum (see here for the classic Blue Tit example). Surely we can ‘play on’ the differences in colour vision between target and non-target species to reduce bycatch and maintain fish catch? Different coloured nets perhaps?
While many seabirds are likely to be able to perceive fine differences in colour across the visible spectrum (violet [400nm] – red [650nm]) in high light conditions, most will not see into the UV and certainly do not detect UV at low light levels. Considering this, and the fact that only a narrow band of wavelengths penetrate to depth, ‘playing on’ colour to reduce bycatch underwater probably isn’t the most fruitful route.
Something else to consider is what a bird is actually looking at. It won’t have escaped your attention that bird’s eyes are on the side of the skull, and not facing forwards. The distribution of photoreceptors (the cells the convert light into signals to send to the brain) on the retina reflect this, meaning that the areas of highest acuity project laterally from the direction of forward travel. In essence, this means birds are not necessarily looking (or at least not looking particularly well) where they are going.
What can be done?
Given all of the above, it is hardly surprising that birds are caught in gillnets in such large numbers. We can conclude that birds do not see gillnets – they forage in low light levels, unable to see fine detail. But what does this mean for how we solve this problem? Mitigation measures need to be developed which:
- Can alert the species to the presence of the net over a wide range of light levels.
- Do not disrupt the dark adapted state of the animal’s retinas.
- Have a high probability of being detectable by all animals that are subject to gillnet bycatch.
- Are relatively simple to deploy and robust under sea conditions.
- Do not reduce the catch of target fish by a significant percentage.
This sounds like a tall order, but Graham has alighted on an idea – stimulus warning panels, attached to nets at regular intervals (every 4 metres). These panels (Fig.2) can be printed on flexible plastic sheet and can be deployed with the net. They are designed to maximise contrast by employing black and white grating or checkerboard patterns. Based on what is known of bird vision underwater, such panels ought to alert birds to the presence of nets from a distance of at least 2 metres in low light conditions. Such panels would also be visible to pinnipeds, sea turtles and larger non-target fish, such as blue water predators including tuna. What we need to understand is the behavioural response of both target fish and non-target species to these panels. Will they meet the 5 criteria listed above, alert birds to the presence of the net and if possible deterring them from the net area, without deterring fish completely? The only way to find out is through field trials – and BirdLife are in the process of investigating this in fisheries in Lithuania and Chile.
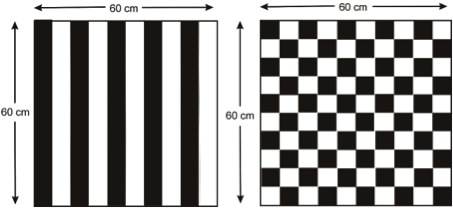
Fig. 2. Examples of a grating and checkerboard patterns of high contrast recommended for use as stimulus warning panels. The overall dimensions are indicated, the dimension of individual elements, stripes widths and sides of squares, are 6 cm.
When this blog is published, Graham and I will be on our way to discuss this idea, hopefully among many others, at a gillnet bycatch workshop BirdLife are co-hosting with the American Bird Conservancy in the USA. Turtle, seabird and cetacean experts from across the world will be attending, alongside fishermen, government staff and sensory ecologists. We’re hopeful the workshop will spawn a number of research projects aimed at reducing the toll of gillnet bycatch on non-target species.
There is a long way to go to solve this problem – but as we take the first steps, we hope we’re heading in the right direction.
References and further reading
Anderson, O.R., Small, C.J., Croxall, J.P., Dunn, E.K., Sullivan, B.J., Yates, O. & Black, A. 2011. Global seabird bycatch in longline fisheries. Endang. Species Res. (14) 91–106.
Croxall, J.P., Butchart, S.H.M., Lascelles, B., Stattersfield, A.J., Sullivan, B., Symes, A. & Taylor, P. 2012. Seabird conservation status, threats and priority actions: a global assessment. Bird Conserv. Int. (22) 1–34.
Martin, G.R & Crawford, R. 2014. Reducing bycatch in gillnets: A sensory ecology perspective. Global Ecology and Conservation (3) 28-50.
White, C.R., Day, N., Butler, P.J., Martin, G.R., 2007. Vision and foraging in cormorants: more like herons than hawks? PLoS ONE 2 (7), e639. https://dx.doi.org/10.1371/journal.pone.0000639.
Zydelis, R., Bellebaum, J., Osterblom, H., Vetemaa, M., Schirmeister, B., Stipniece, A., Dagys, M., van Eerden, M. & Garthe, S. 2009. Bycatch in gillnet fisheries – an overlooked threat to waterbird populations. Biol. Conserv. (142) 1269–1281.
Zydelis, R., Small, C. & French, G. 2013. The incidental catch of seabirds in gillnet fisheries: a global review. Biol. Conserv. (162) 76–88.
Image credits
Top: Steller’s Eider in net © Markus Vetemaa
If you want to write about your research in #theBOUblog, then please see here.

