

 LINKED PAPER
LINKED PAPER
Nest site selection and reproductive success of a critically endangered parrot, the great green macaw (Ara ambiguus), in an anthropogenic landscape. Lewis, T.C., Gutiérrez Vargas, I., Vredenbregt, C., Jimenez M., Hatchwell, B., Beckerman, A.P., Childs, D.Z. 2023 Ibis. doi: 10.1111/ibi.13262 VIEW
Globally, there are 1307 secondary cavity-nesting bird species, of which 13% are threatened with extinction (van der Hoek et al. 2017). These species rely on cavities created by decay and other species to nest in. Nest site selection is the principal way secondary cavity-nesting species mitigate the negative effects of factors such as predation, parasitism and exposure on reproductive success. Large-bodied secondary cavity nesters, like the Great Green Macaw (Ara ambiguus), are particularly reliant on mature trees that contain cavities of sufficient size (Marsden and Jones 1997; de la Parra-Martínez et al. 2015; Renton et al. 2015). However, large cavities in mature trees are often absent or reduced in anthropogenically disturbed forests. Thus, the availability of high-quality nest sites may be limited for these species, potentially reducing reproductive success.
Our study area is a section of the San Juan La Selva biological corridor in northern Costa Rica. It is a highly anthropogenically disturbed landscape with a mix of cattle pasture, pineapple plantations and primary and secondary forest. Although forest cover in the area has increased in the last 20 years, there are still ongoing pressures from logging and pineapple expansion, driving habitat degradation.
To investigate how GGMs select nest sites in this area, we compared the features of active nests with unused cavities found within a 500m radius of the active nests. We used a drone to identify potential cavities in emergent trees and then climbed to and measured these cavities by hand. Due to the pandemic, we didn’t have time to climb ~80 trees with cavities, meaning we missed a potential 220 extra cavities.
At the same time, the team monitored breeding attempts at 33 active nests. These known nest sites come from a database managed by the Centro Cientifico Tropical and Macaw Recovery Network that covers known nest sites from 1995 to the present. We used the number of fledglings, which we defined as the number of fully feathered chicks last seen in the nest before identifying an empty nest, as our reproductive metric.
To investigate nest-site selection, we fitted a pair of intercept-only Bayesian regression models for occupied and unoccupied for each feature. We used the posterior distribution of the means and standard deviations to summarise their uncertainty and evaluate whether GGMs selected for a cavity feature. This showed that GGMs select for cavity depth, internal diameter, entrance height, entrance size and canopy connectivity.
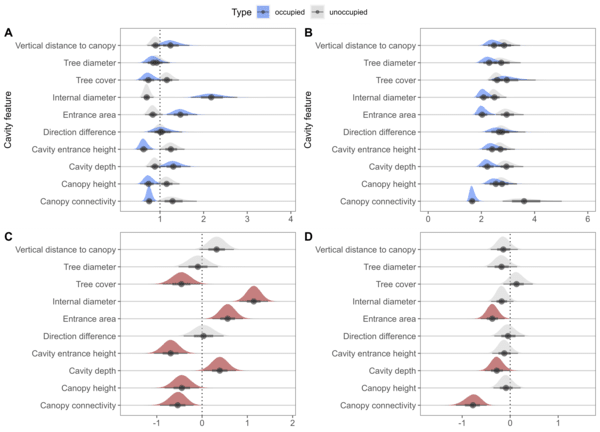
Figure 1 Univariate model A) means and posterior distributions and B) variance and posterior distributions for occupied and unoccupied cavities. With the difference in C) means and posterior distributions and D) standard deviation and posterior distributions for each cavity feature with significant features, those with 95% credibility intervals not crossing zero, are highlighted in red.
We wanted to create a single score for a cavity that would indicate its suitability. So, for each feature, we calculated the number of standard deviations away from the model mean. We then took the mean of all these differences in standard deviation for each cavity to create the nest suitability score.
Overall, 1.34 fledglings were produced per breeding attempt. This is within the range of other large macaws. We were able to attribute five failures to predation; four of the failures were attributed to avian predators. Interestingly, during the previous breeding season, there were three confirmed predation events. One was attributed to avian predators; this nest was also predated in 2020.
We then carried out an ordinal regression for each feature, using the nest suitability score and the selected features as individual predictor variables, with reproductive output as the response variable. Cavity depth and nest suitability score were the only two that were significantly positively correlated with reproductive success.
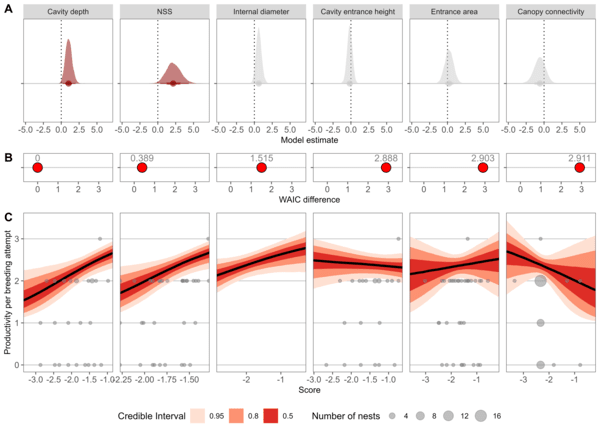
Figure 2 The relationship between productivity and cavity features. A) Model estimates of the regression slopes and their posterior distributions from the univariate ordinal regression of each cavity feature, with the significant models in red. B) The difference in WAIC scores compared to the best-performing model. C) The relationship between each feature and predicted productivity alongside the data (grey).
Finally, we determined how many high-quality cavities were available – those with predicted productivity higher than the mean reproductive success. The number of high-quality cavities available varies depending on which metric is used. There were 39 high-quality available cavities when using cavity depth and only 17 high-quality cavities when using the nest suitability score. However, interspecific competitors use 20% (cavity depth) and 10% (nest suitability score) of these.
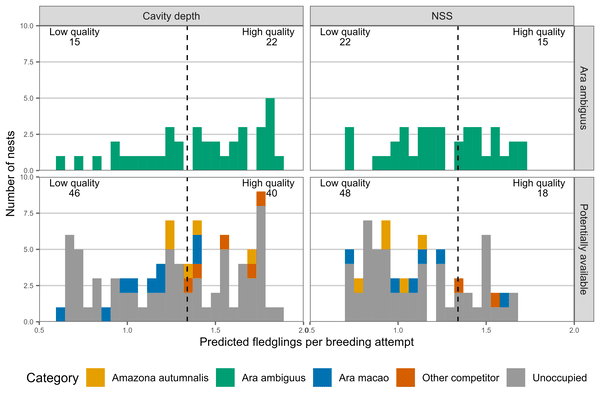
Figure 3 Comparison of occupied and unoccupied cavity features. Distribution of the predicted number of fledglings per breeding attempt from our univariate models. We restricted the unoccupied cavities to the range of the occupied cavities in both cases. The dashed line is the mean number of fledglings per breeding attempt from our monitoring data. The cavities to the right of the mean line are defined as high-quality and to the left, as low-quality.
Great Green Macaws in our study population prefer larger, deeper cavities in isolated trees, showing a strong selection for larger cavity entrance area, greater depth and low canopy connectivity. High-quality nest sites are those that are deep. Our combined nest suitability score is also a good metric of nest quality, suggesting that although not significant on their own, other cavity features also contribute to reproductive success. The selection of these features is likely a mechanism to reduce the risk of predation
Great Green Macaws also select for nests that are low in trees. This makes sense for big birds, as lower in the tree is more able to accommodate large cavities. However, some nest entrances in open areas were only 9m and 10m off the ground! The average canopy height in the region is 24m. This presents a challenge; if the forest regenerates around these nests, they will become unusable, further reducing the number of available cavities. Trees with cavities large enough to accommodate Great Green Macaws take a long time to grow. We estimate the youngest of the primary nest tree species, Mountain Almond Dipteryx oleifera, is nearly 300y/o! Solution?
- Protect all trees so we give nature as much opportunity as possible to create new cavities
- While we wait, we can provide nest boxes near those nests that are being covered by regenerating forest
Nest-box provision could be useful as they could be designed to reduce risk from avian predation. However, there is also a long-term commitment needed to their maintenance.
References
de la Parra-Martínez, S.M., Renton, K., Salinas-Melgoza, A., Muñoz-Lacy, L.G. 2015. Tree cavity availability and selection by a large-bodied secondary cavity-nester: the Military Macaw. Journal of Ornithology 156:489-498. VIEW
Marsden, S.J., Jones, M.J. 1997. The nesting requirements of the parrots and hornbill of Sumba, Indonesia. Biological Conservation 82:279-287. VIEW
Renton, K., Salinas-Melgoza, A., De Labra-Hernández, M.Á., de la Parra-Martínez, S.M. 2015. Resource requirements of parrots: nest site selectivity and dietary plasticity of
Psittaciformes. Journal Ornithology 156:73-90. VIEW
van der Hoek, Y., Gaona, G.V., Martin, K. 2017. The diversity, distribution and conservation status of the tree-cavity-nesting birds of the world. Diversity Distributions 23:1120-1131. VIEW
Image credit
Top right: A pair of Great Green Macaws perched on the edge of their nest cavity in northern Costa Rica © Corey Raffel.
If you want to write about your research in #theBOUblog, then please see here.

