

 European farmland areas are undergoing immense change, particularly in uplands habitats. In recognition of the severe biodiversity declines in farmland areas related to farming intensification, a variety of agri-environmental schemes have been implemented, with mixed results (Ekroos et al. 2014). Monitoring the success of these schemes is particularly challenging and labour intensive. There is an urgent need for reliable indicators for rapid assessments of the effect of changes in management practices and for restoration/rehabilitation projects. Since birds have diverse roles within food webs, avian soundscapes may provide such an indicator, allowing for passive, cost-effective monitoring of the biological outcomes of management changes and restoration.
European farmland areas are undergoing immense change, particularly in uplands habitats. In recognition of the severe biodiversity declines in farmland areas related to farming intensification, a variety of agri-environmental schemes have been implemented, with mixed results (Ekroos et al. 2014). Monitoring the success of these schemes is particularly challenging and labour intensive. There is an urgent need for reliable indicators for rapid assessments of the effect of changes in management practices and for restoration/rehabilitation projects. Since birds have diverse roles within food webs, avian soundscapes may provide such an indicator, allowing for passive, cost-effective monitoring of the biological outcomes of management changes and restoration.
A lack of rapid assessment tools for biodiversity often constrains the progress of remedial measures and crucially, the evaluation of habitat management, restoration, and determination of best practice. This is particularly the case where data is required to direct initiatives promoting changes in management practices, such as in European farmlands. Long-term experimental systems are a rare but powerful way to evaluate changes in management practices.
Passive acoustic monitoring (PAM) is emerging as a important tool for inferring biodiversity and ecosystem function, as diverse ecosystems should exhibit complex acoustic signals (Ross et al. 2023). Analysis of soundscapes can be time- and cost-effective, compared to traditional methods (Darras et al. 2019). Since birds have diverse roles within food webs, their diversity and community structure can be used to evaluate the outcomes of habitat management (Wenny et al. 2011; Canales-Delgadillo et al. 2019).
In this study we tested whether passive acoustic monitoring can be used to detect avian biodiversity change in the long-term Glen Finglas livestock-grazing experiment. The Glen Finglas experiment in central Scotland (56°16ʹ03ʺN, 4°25ʹ08ʺW), has been running continuously since 2003. In this experiment 3.3. ha. plots have been managed with different set grazing intensities, or left ungrazed, allowing assessment of the effect of management and restoration on biodiversity and ecosystem function (Evans et al. 2005). Recent work has shown an increase in bird species richness in the ungrazed plots, driven by changes in vegetation (Malm et al. 2020).

Figure 1 The difference between grazed and ungrazed plots © Robin Pakeman.
To assess whether PAM could be used in continued long-term monitoring of avian biodiversity trends in this upland system, requiring less intensive survey effort than previous work (Malm et al. 2020), we placed PAM sensors in paired experimental plots. Six PAM sensors were placed in plots which were grazed at a commercial stocking density of sheep with nine ewes per plot, and six PAM sensors were placed in ungrazed plots. Sound recording was undertaken 24 hours of the day, with recording carried out for two minutes in every 10 minute interval. Recording was undertaken between the start of April and June 2022, focusing on the peak of bird acoustic activity, during territory formation and breeding, producing 130,000 minutes of soundscape recordings, in individual two minute audio files. For each individual audio file, we calculated a series of standard acoustic indices including Acoustic Complexity Index (ACI), Acoustic Diversity Index (ADI), Acoustic Evenness (AEve), Bioacoustic Index (Bio), Acoustic Entropy (H) and Normalized Soundscape Index (NDSI), which can provide insight into different aspects of biodiversity and soundscape properties (Bradfer-Lawrence et al. 2019). For each of these acoustic indices, we analysed whether they were significantly different between ungrazed and intensively grazed plots. The analyses were carried out for the full 24 hour span, and for four focal hours in the day separately; the hour around dawn, dusk, midday and midnight, to account for daily temporal variability in the soundscape. Additionally, to account for phenological variability in the soundscape, we carried out our analysis on the full time of recording, then for April and May separately.

Figure 2 SM4 acoustic recorder in place in a study plot at Glen Finglas. Chicken wire was used to protect the recorders from sheep in the intensively grazed plots. © Darren Evans.
The highlight result of our analyses was that the Bioacoustic Index was significantly higher during the dawn chorus in ungrazed plots, though only in April. This index did not differ between treatments at other times of the day. The Bioacoustic Index was developed to monitor avian biodiversity, with higher values indicating higher avian biodiversity (Boelman et al. 2007). Therefore, our result suggests that there higher avian biodiversity in the ungrazed plots than in the intensively grazed plots, confirming our hypothesis. This result indicates that it is possible to track avian biodiversity change via PAM, even in relatively low diversity upland systems. It also makes clear the importance of targeting biologically relevant times of day, and of year, informed by the study group’s phenology. At this relatively high latitude, it is likely that there is a greater intensity of vocalisations in April, earlier in the breeding season, as territory boundaries are solidified, and before nestlings hatch focusing parental resources more on provisioning. Additionally, as expected, vocalisation effort appeared focused during the dawn chorus period, which was the only time of day where a significant difference was found between ungrazed and intensively grazed plots.
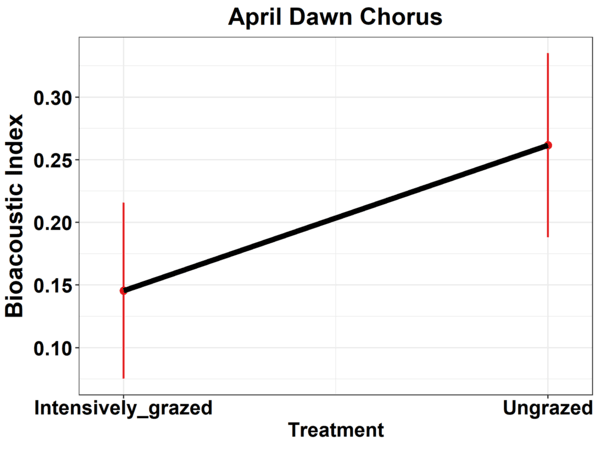
Figure 3 The impact of plot treatment (ungrazed versus intensively grazed plots) on the Bioacoustic Index derived from acoustic monitoring. Modelled relationship ±95% confidence intervals displayed © Darren O’Connell.
Following on from this, our next steps will be to focus on the individual species patterns that may be driving change in the acoustic indices, using machine learning techniques to identify individual species in the soundscape (Ross et al. 2018). This will allow us to assess how different species may be responding the change in land use in terms of both abundance and phenology, with a particular focus on how key upland indicator species such as the Meadow Pipit (Anthus pratensis). We hope this will provide a template for monitoring recovering habitats in all ecosystems.
Acknowledgements
Darren Evans of Newcastle University carried out all fieldwork for this project and Samuel Ross of the Okinawa Institute of Science and Technology provided support to the project design. This project was supported by a British Ornithologists’ Union Small Ornithological Research Grant and Wildlife Acoustics Scientific Product Grant (2019Q4). The Glen Finglas grazing experiment has been run since 2003 by the James Hutton Institute. This has been facilitated by the Woodland Trust, who are the landowners and have been conducting wider restoration activities on the Glen Finglas property.
References
Boelman, N.T., Asner, G.P., Hart, P.J., Martin, R.E. 2007. Multi-trophic invasion resistance in Hawaii: bioacoustics, field surveys, and airborne remote sensing. Ecological Applications 17:2137-2144. VIEW
Bradfer-Lawrence, T, Gardner, N, Bunnefeld, L, Bunnefeld, N, Willis, SG, Dent, DH. 2019. Guidelines for the use of acoustic indices in environmental research. Methods in Ecology and Evolution 10:1796-1807. VIEW
Canales-Delgadillo, JC, Perez-Ceballos, R, Zaldivar-Jimenez, MA, Merino-Ibarra, M, Cardoza, G, and Cardoso-Mohedano, JG. 2019. The effect of mangrove restoration on avian assemblages of a coastal lagoon in southern Mexico. PeerJ 7:e7493. VIEW
Darras, K, Batáry, P, Furnas, BJ, Grass, I, Mulyani, Y A, and Tscharntke, T. 2019. Autonomous sound recording outperforms human observation for sampling birds: a systematic map and user guide. Ecological Applications 29:e01954. VIEW
Ekroos, J, Olsson, O, Rundlöf, M, Wätzold, F, and Smith, HG. 2014. Optimizing agri-environment schemes for biodiversity, ecosystem services or both?. Biological Conservation 172:65-71. VIEW
Evans, DM, Redpath, SM, Evans, SA, Elston, DA and Dennis, P. 2005. Livestock grazing affects the egg size of an insectivorous passerine. Biology Letters 1:322-325. VIEW
Malm, LE, Pearce‐Higgins, JW, Littlewood, NA, Karley, AJ, Karaszewska, E, Jaques, R, Pakeman, RJ, Redpath, SM and Evans, DM. 2020. Livestock grazing impacts components of the breeding productivity of a common upland insectivorous passerine: Results from a long‐term experiment. Journal of Applied Ecology 57:1514-1523 VIEW
Ross SRP-J, Friedman NR, Dudley KL, Yoshimura M, Yoshida T, and Economo EP. 2018. Listening to ecosystems: data-rich acoustic monitoring through land-scape-scale sensor networks. Ecological Research 33:135-147. VIEW
Ross, SRP-J, O’Connell, DP, Deichmann, JL, Desjonquères, C, Gasc, A, Phillips, JN, Sethi, SS, Wood, CM, and Burivalova, Z. 2023. Passive acoustic monitoring provides a fresh perspective on fundamental ecological questions. Functional Ecology 37:959-975. VIEW
Wenny DG, De Vault TL, Johnson MD, Kelly D, Sekercioglu CH, Tomback DF, and Whelan CJ. 2011. The need to quantify ecosystem services provided by birds. The Auk 128:1-14. VIEW
Image credit
Top right: Meadow Pipit © Gerry Zambonini by CC BY-SA 3.0 Wikimedia Commons.

