

 LINKED PAPER
LINKED PAPER
Novel food resources and conservation of ecological interactions between the Andean Araucaria and the Austral parakeet. Blanco, G., Romero‐Vidal, P., Tella, J. L., & Hiraldo, F. 2022 Ecology and Evolution. doi: 10.1002/ece3.9455 VIEW
The Austral Monkey Puzzle (Araucaria araucana; hereafter Araucaria) constitutes a keystone species in the highly seasonal Southern Andean Forest (Tella et al. 2016). However, this species is under threat as a result of several anthropogenic activities, such as overexploitation of timber, agriculture and livestock farming (Premoli et al. 2013). As a consequence, the Araucaria forest has lost up to 40% of its usual distribution, and has led to the species being classified as Endangered by the IUCN.
The Araucaria has an interesting masting seed production strategy (Sanguinetti & Kitzberger 2008). This plant produces a large number of seeds in some years alternating with others of lower production, thus differentiating between masting and non-masting years. Producing temporally concentrated high quantities of large seeds during masting years, allows the plant to satiate predators that act as the main seed dispersers.
The Austral Parakeet (Enicognathus ferrugineous) has developed a close ecological and evolutionary relationship with the Araucaria. These parrots act as pollinators of the female cones (Gleiser et al. 2019), and consume and disperse large quantities of seeds, being considered as the most important long-distance disperser for this species (Tella et al. 2016). However, nowadays Araucaria seeds are also consumed by a wide variety of human introduced mammals, including livestock (Tella et al. 2016). In fact, the large number of seeds consumed during non-masting years could be depleting this resource, thus displacing the Austral Parakeet and altering the ecological interaction between these two species.

Figure 1 Austral Parakeets feeding on (a) Araucaria seeds and (b) pollen from Araucaria male cones in native forests, and fruits of exotic (c) Malus domestica and (d) Cotoneaster sp. in urban areas of the Northern Patagonian Andes © Orlando Mastrantuoni.
To assess the alteration in the interactions between Araucaria and Austral Parakeets due to exotic mammals, we designed a sampling study in northwestern Patagonia, comprising Argentina and Chile. The study area included both Araucaria Forested areas and areas with different degrees of transformation, comprising mosaic landscapes including Nothofagus Forest, Araucaria Forest, steppe-scrubland, urban and agricultural land cover types. In the selected area, we conducted roadside surveys to estimate Austral Parakeet abundances. We choose this methodology since it has been shown as a reliable approach for estimating abundances of highly mobile, gregarious species with patchy distributions, such as psittacine species (Tella et al. 2021). We performed these censuses during the Austral Parakeet’s breeding and non-breeding seasons, from 2013 to 2016. We also recorded data on the habitat type where parakeets were counted, considering them as habitat patches, and each one of them as a sampling unit.
When Austral Parakeets were detected feeding we stopped to record the following data: 1) flock size, 2) plant species, 3) item exploited (only in Araucaria) and 4) habitat. Plants consumed by Austral Parakeets were also categorized as either native or exotic. Finally, we measured seed abundance below the same marked Araucaria trees during masting and non-masting years, and during breeding and non-breeding seasons.
Our surveys that were performed across more than 12,000km over four years showed that in general, parakeets occurred in patchy distributions, with higher relative abundances in the non-breeding season. The results suggested differences between habitats depending on masting and non-masting years.
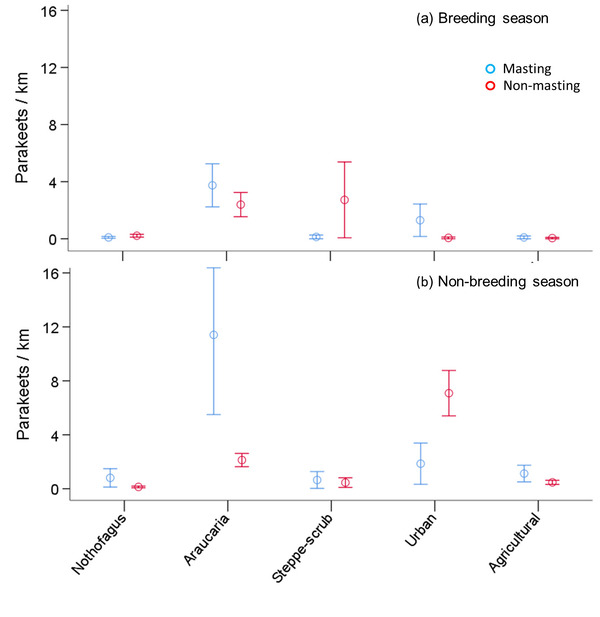
Figure 2 Relative abundance of Austral Parakeets (mean ± SE individuals per km) recorded during roadside car surveys in each habitat type according to Araucaria crop abundance (masting or non-masting) during the (a) breeding season and (b) non-breeding season in the Northern Patagonian Andes.
When looking to feeding patterns, Austral Parakeets fed in all habitats. However, we observed a higher proportion of flocks exploiting native, pure and mixed Araucaria and Nothofagus Forests (native species forests) during the breeding season, especially during masting years, while in non-masting years usage of urban areas increased considerably. Native habitat use by Austral Parakeets was higher in masting years (91.1%) than in the non-masting years (69.4%) during the breeding season. This difference in habitat use was more acute during the non-breeding season, with 85.7% of flocks occurring in native habitats during masting years, and only 34.2% during non-masting years.
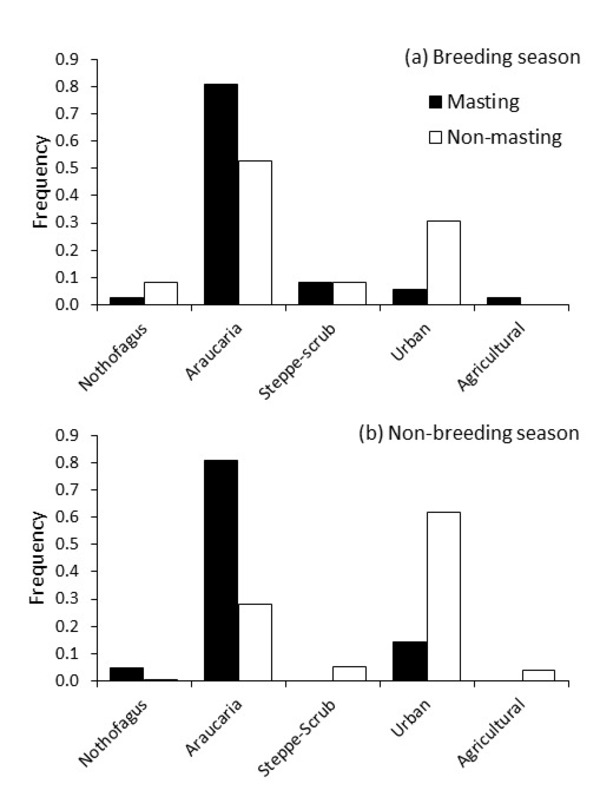
Figure 3 Frequency of Austral Parakeet flocks (n = 354, including single individuals) foraging in each habitat type according to Araucaria crop abundance (masting or non-masting) during the (a) breeding season and (b) non-breeding season in the Northern Patagonian Andes.
Of the 354 flocks observed feeding, totalling nearly 20,000 individuals, we were able to identify 18 native and 21 exotic plant species on which they fed. Parakeets consumed a wide variety of plant food items, including Araucaria seeds, pollen and sap, as well as seeds, fruits, flowers, flower buds, leaves, leaf buds, sap, bark and twigs of native and exotic plants, including wild herbs and cultivated cereal grain, fruit trees and garden plants in urban areas.
About half of foraging flocks were observed feeding on native species, while the other half fed on exotic species. However, exploitation of exotic plants by flocks was lower in masting years (93.1% native, 6.9% exotic) than in non-masting years (39.5% native, 60.5% exotic). Foraging flock size was also influenced by masting and non-masting years as well as food type (native and exotic). During the breeding season, we detected smaller flocks than in the non-breeding season and flock size was larger in masting years. Finally, larger flocks exploited native plants during masting years in the non-breeding season and were even larger when exploiting exotic plants in non-masting years.
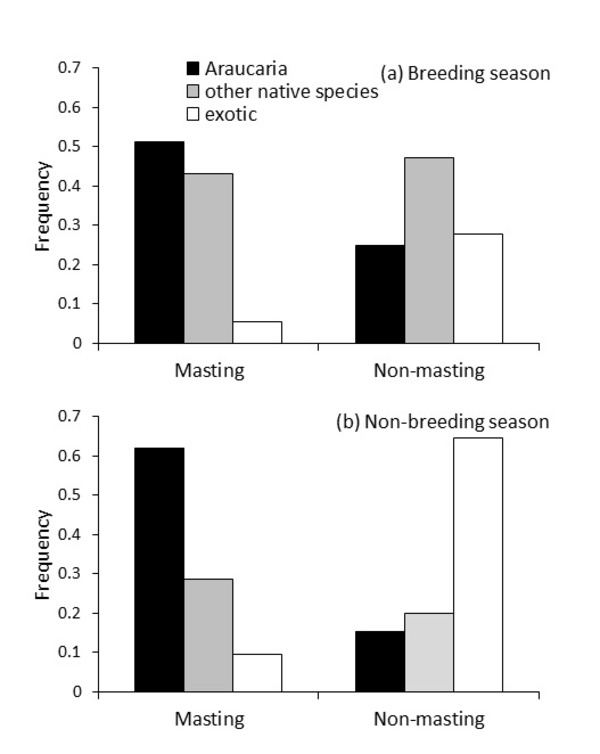
Figure 4 Frequency of Austral Parakeet flocks (n = 354, including singe individuals) foraging on Araucaria, other native plant species, and exotic species according to Araucaria crop abundance (masting or non-masting) during the (a) breeding season and (b) non-breeding season in the Northern Patagonian Andes.
The results of this study show that the masting seed production strategy of Araucaria influences the inter-annual dependence of Austral Parakeets in native habitats. We observed how during non-masting years, Austral Parakeets shifted to exotic plants in anthropogenic habitats. The use of anthropogenic habitats during masting seed production years was residual, indicating a clear preference for the Araucaria seeds if this resource was available. However, during non-masting years flocks shifted to exotic plants which was also reflected by a larger flock size in anthropogenic habitats during these years.
The nomadic and migratory strategies for the exploitation of resources in the highly seasonal environment of the Patagonian Andes could allow Austral Parakeets to switch to novel resources exploited in anthropogenic habitats depending on the availability of native resources, especially Araucaria seeds. The reduction of the Araucaria Forest, coupled with the consumption of seeds by exotic mammals in non-masting years is forcing Austral Parakeets to seek new food sources in anthropogenic habitats. This not only leads to the loss of ecological interactions between these two species but may also pose a conservation problem for Austral Parakeets. Feeding on crops increases the negative perception of this species, reinforcing parrot-human conflict that is already occurring in other parts of the Neotropics
References
Sanguinetti, J., & Kitzberger, T. 2008. Patterns and mechanisms of masting in the large-seeded southern hemisphere conifer Araucaria araucana. Austral Ecology 33: 78-87. VIEW
Tella, J. L., Lambertucci, S. A., Speziale, K. L., & Hiraldo, F. 2016. Large-scale impacts of multiple co-occurring invaders on monkey puzzle forest regeneration, native seed predators and their ecological interactions. Global Ecology and Conservation 6: 1-15. VIEW
Premoli, A. C., Quiroga, P., & Gardener, M. R. 2013. Araucaria araucana. The IUCN Red list of threatened species 2013. IUCN.
Gleiser, G., Speziale, K. L., Lambertucci, S. A., Hiraldo, F., Tella, J. L., & Aizen, M. A. 2019. Uncoupled evolution of male and female cone sizes in an ancient conifer lineage. Journal of Plant Sciences 180: 72-80. VIEW
Tella, J.L., Romero-Vidal, P., Dénes, F.V., Hiraldo, F., Toledo, B., Rossetto, F., Blanco, G., Hernández-Brito, D., Pacífico, E., Díaz-Luque, J.A., Rojas, A., Bermúdez-Cavero, A., Luna, Á., Barbosa, J.M. & Carrete, M. 2021. Roadside car surveys: methodological constraints and solutions for estimating parrot abundances across the world. Diversity 13: 300. VIEW
Image credit
Top right: Austral Parakeet Enicognathus ferrugineous © José Luis Tella.
If you want to write about your research in #theBOUblog, then please see here.

