

Report from a BOU-funded project
LINKED PAPER
No sex-specific differences in the influence of high air temperatures during early development on nestling mass and fledgling survival in the Southern Pied Babbler (Turdoides bicolor) Bourne, A.R., Cunningham, S.J., Nupen, L.J., Mckechnie, A.E. & Ridley, A.R. 2021 IBIS. DOI: 10.1111/ibi.12990 VIEW
Researchers at the Pied Babbler Research Project have been studying breeding behaviour in Southern Pied Babblers Turdoides bicolor since 2003 (Ridley 2016). Southern Pied Babblers (60-90g) are medium-sized, cooperatively breeding passerines found in the semi-arid savannas of southern Africa’s Kalahari (Ridley 2016). They are highly monomorphic, which means that males and females cannot readily be distinguished from one another by the way that they look or behave.
 Figure 1: A family of Southern Pied Babblers take some time out to preen each other © Marc Freeman
Figure 1: A family of Southern Pied Babblers take some time out to preen each other © Marc Freeman
The team has found that pied babblers aren’t able to gain enough mass from foraging to counter typical overnight mass loss (du Plessis et al. 2012) on days when the maximum temperature exceeds 35.5 °C. Breeding babblers also substantially reduce the amount they feed their young when this threshold is crossed (Wiley & Ridley 2016). When temperatures have been very high, nestlings measured at 11-days old are smaller as a result (Wiley & Ridley 2016). Whether there are sex-specific differences in the impacts of extreme heat has not previously been studied in pied babblers.
In lots of different species, including birds and humans, male young are more vulnerable to death during early development than female young. There are several theories to explain this (Sheldon et al. 1998): 1) the sex carrying two chromosomes (e.g. in humans, females are XX and males XY) will be weaker, 2) in species where one sex is larger as an adult, young of the larger sex will grow larger and faster and need more food to be as viable as the smaller sex, and 3) the energy demands of higher levels of testosterone in males compromises immune function.
 Figure 2: As part of the fieldwork for this research we ring, measure, and blood sample nestlings when they are 11 days old © Todd Erickson
Figure 2: As part of the fieldwork for this research we ring, measure, and blood sample nestlings when they are 11 days old © Todd Erickson
In birds, females carry the extra chromosome – females are ZW and males are ZZ – so this cannot explain male-biased mortality in birds. The second theory is well supported, with many species showing higher juvenile male mortality where males are the larger sex. However, this effect is still evident in size-monomorphic species, and there is no evidence that the trend is reversed in species where females tend to be larger (Clutton-Brock et al. 1985). Studying sex-specific differences in a monomorphic bird is interesting because any evidence of male-biased mortality would suggest that there is a physiological basis for this, supporting theory 3. We used long term nestling life history data to test for any sex-specific differences in size, mass, and survival of pied babbler nestlings exposed to high temperatures during early development.
There is a small but consistent female-skew in sex ratios at various stages during early development (Ridley 2016). Around 46% of 11-day old nestlings are male, suggesting higher male mortality. This slight female bias is maintained in babblers that survive to 90 days of age (when young are considered nutritionally independent – Ridley 2016). Approximately 45% of independent juveniles are male. We measured the effect of sex, temperature, and rainfall on body mass in 368 nestlings, tarsus length in 217 nestlings, and survival to independence in 551 nestlings.
Pied babbler nestlings at 11-days old are negatively affected by high temperatures, being both lighter (by body mass in grams) and smaller (by tarsus length in mm) when there has been a high proportion of days during the nestling period above the critical temperature threshold of 35.5 °C. However, there was no difference between male and female nestlings in terms of either body mass or tarsus length, and maximum air temperature affected male and female nestlings in the same way (Fig. 3).
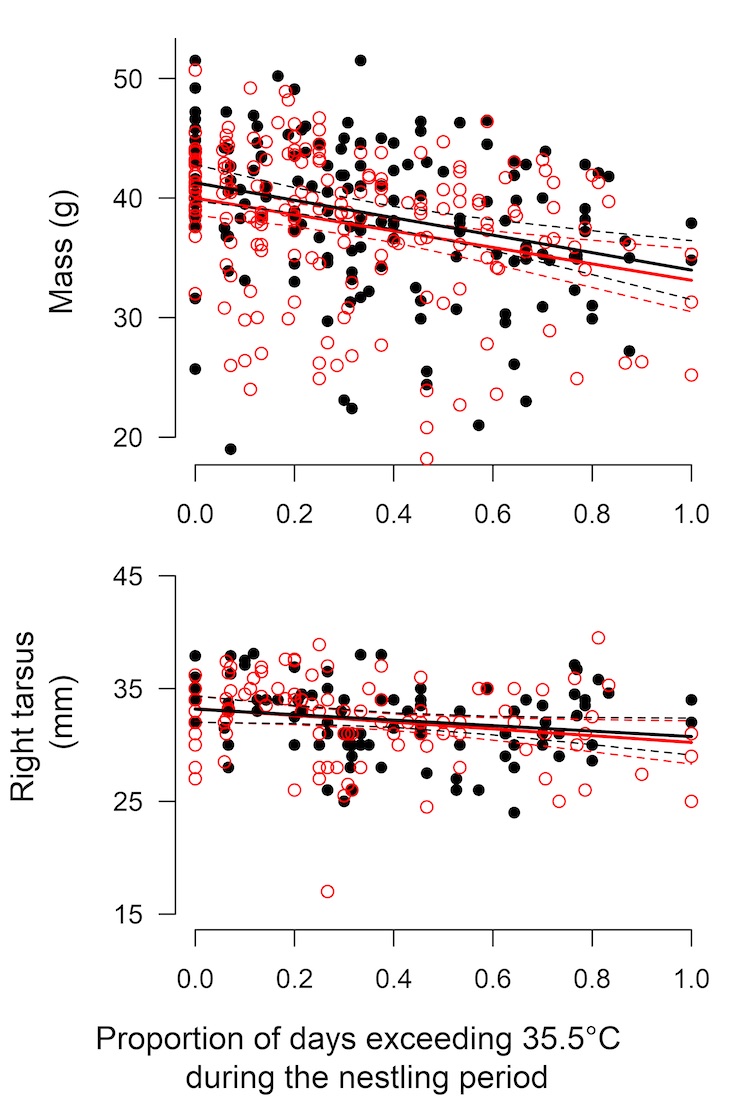 Figure 3: Southern Pied Babbler Turdoides bicolor nestlings are affected by high temperatures in the nest – with nestlings both weighing less and having shorter tarsi at 11 days old at high proportions of hot days during the nestling period. This effect of temperature does not differ between male (black filled circles and black lines) and female (red open circles and red lines) nestlings. Solid lines represent the regression output and dashed lines the 95% confidence interval.
Figure 3: Southern Pied Babbler Turdoides bicolor nestlings are affected by high temperatures in the nest – with nestlings both weighing less and having shorter tarsi at 11 days old at high proportions of hot days during the nestling period. This effect of temperature does not differ between male (black filled circles and black lines) and female (red open circles and red lines) nestlings. Solid lines represent the regression output and dashed lines the 95% confidence interval.
Contributors: Amanda Ridley, Susan Cunningham.
Approximately 60% of all nestlings in our study population survive to independence. Heavier nestlings of both sexes were more likely to survive to independence, as were those fledglings that enjoyed higher rainfall between fledging and independence (presumably because there is more food available when conditions are wetter). We found that high temperatures after leaving the nest reduced the chances of surviving to independence slightly (the effect is not statistically significant) and that male and female nestlings were equally likely to survive to independence (Fig. 4).
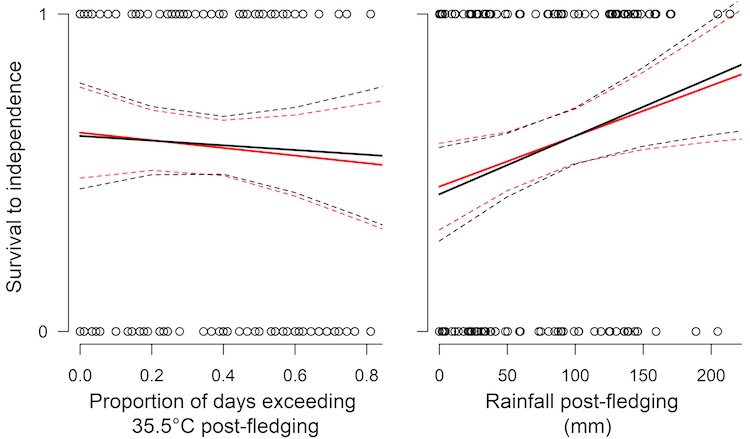 Figure 4: Juvenile survival to independence (90 days after fledging) in Southern Pied Babblers Turdoides bicolor is affected by rainfall but not by temperature after leaving the nest. Effects do not differ between male (black lines) and female (red lines) nestlings. Solid lines represent the regression line and dashed lines the 95% confidence interval.
Figure 4: Juvenile survival to independence (90 days after fledging) in Southern Pied Babblers Turdoides bicolor is affected by rainfall but not by temperature after leaving the nest. Effects do not differ between male (black lines) and female (red lines) nestlings. Solid lines represent the regression line and dashed lines the 95% confidence interval.
Contributors: Amanda Ridley, Susan Cunningham.
As may be expected for a highly monomorphic species, we found no evidence for sex-specific responses to high temperatures in terms of nestling growth and survival. While the cause of the consistent female bias in the sex ratio of nestlings and juvenile pied babblers remains something of a mystery, we found plenty of evidence for significant climate-related impacts on nestling development and survival, and we continue to investigate both of these phenomena.
Acknowledgments
The project received a research grant of £1,532 from the BOU in 2017 for which the project team is extremely grateful. The author wishes to thank her supervisors Dr Susan Cunningham and Associate Professor Amanda Ridley, her field assistant Mr Sello Matjee, and numerous staff and volunteers at the Kalahari Research Centre, Kuruman River Reserve, for their support and assistance with fieldwork. This research received additional funding from grants to Dr Susan Cunningham from the National Research Foundation, South Africa, the University of Cape Town, and the DST-NRF Centre of Excellence at the FitzPatrick Institute of African Ornithology.
References
Clutton-Brock, T. H., Albon, S. D. & Guiness, F. E. 1985. Parental investment and sex differences in juvenile mortality in birds and mammals. Nature 313:131–133.
Du Plessis, K. L., Martin, R. O., Hockey, P. A. R., Cunningham, S. J. & Ridley, A. R. 2012. The costs of keeping cool in a warming world: Implications of high temperatures for foraging, thermoregulation and body condition of an arid-zone bird. Global Change Biology 18:3063–3070.
Ridley, A. R. 2016. Southern pied babblers: The dynamics of conflict and cooperation in a group-living society. Pp. 115–132 in Dickinson, J. L. & Koenig, W. (eds.). Cooperative Breeding in Vertebrates: Studies of Ecology, Evolution, and Behavior. Cambridge University Press, Cambridge.
Sheldon, B. C., Merila, J., Lindgren, G. & Ellegren, H. 1998. Gender and Environmental Sensitivity in Nestling Collared Flycatchers. Ecology 79:1939–1948.
Wiley, E. M. & Ridley, A. R. 2016. The effects of temperature on offspring provisioning in a cooperative breeder. Animal Behaviour 117:187–195.
Image credits
Featured image: Southern Pied Babbler Turdoides bicolor © Samantha Kirves

