


Morphology and songs defining what is intra/inter specific variation
LINKED PAPER
Geographic variation and taxonomy of the Arremon taciturnus (Hermann, 1783) species complex (Aves: Passerellidae). Buainain, N., Assis, C. P. & Raposo, M. A. 2017. Journal of Ornithology. DOI: 10.1007/s10336-016-1421-8. VIEW
Birds are thought to be one of the animal groups we know the best, especially when referring to diversity, e.g. the number of bird species that exist in the world. Although this is probably true, it gives us a false idea that we are close to the “real” number of extant bird species, or even that we understand global bird diversity quite well. Traditional estimates suggest we have roughly between 9,000 and 10,000 bird species globally. However, a recent evaluation showed that we may be pretty far from the “real” number, and it could be that about twice as many bird species exist in reality (Barrowclough et al. 2016). Similar results are often have been shown by recent taxonomic studies; especially the ones focused on the Neotropical region, where diversification patterns are much more complex.
Complex patterns of speciation can generate complex patterns of variation, which can be exhibited in morphology, vocalization or distribution of species. Thus, distinguishing what is intra- or inter-specific variations remains part of the taxonomic challenge of Neotropical birds. This of course, poses a big obstacle for conservation biology, because we can’t target conservation strategies to protect species if we don’t know precisely what and where are the species that we are in fact trying to protect.
On the other hand, with technological and methodological advances, including the emergence of molecular methods, modern methods of morphological analyses, and also modern tools and online libraries that allow for sophisticated vocalization analyses, we have significantly improved our capacity to understand intra- or inter-specific variations and thus we can today delimit species much more effectively. However, we are still only beginning to understand such patterns and most species still haven’t been examined under modern tools and concepts.
This is the context of the case of the Pectoral Sparrow, Arremon taciturnus, species-complex. Pectoral Sparrows are small songbirds that inhabit various Neotropical vegetation types in South America, such as humid forests on the east base of the Andes, the Amazon, the Cerrado, the Atlantic forest and dry scrubs in the Caatinga (Fig. 1).
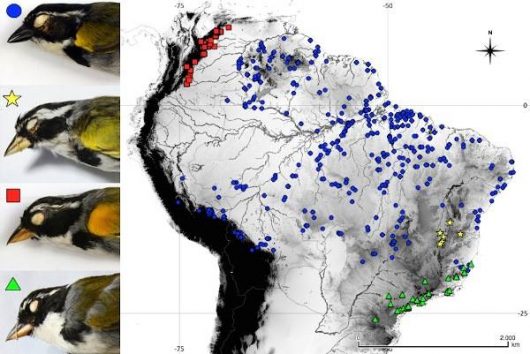 Figure 1 Geographic distribution of the different species of the Pectoral Sparrow, Arremon taciturnus, species complex: A. taciturnus (blue circles), A. franciscanus (yellow stars), A. axillaris (red squares), A. semitorquatus (green triangles). Note the morphological differences between taxa, such as the extent of superciliary stripes and the coloration patterns of the bill and wing coverts
Figure 1 Geographic distribution of the different species of the Pectoral Sparrow, Arremon taciturnus, species complex: A. taciturnus (blue circles), A. franciscanus (yellow stars), A. axillaris (red squares), A. semitorquatus (green triangles). Note the morphological differences between taxa, such as the extent of superciliary stripes and the coloration patterns of the bill and wing coverts
Historically, A. taciturnus is considered a species-complex with A. semitorquatus and A. franciscanus due to their overall morphological similarities. Furthermore, A. taciturnus have two associated subspecies (A. t. axillaris and A. t. nigrirostris), which were originally described as distinct species, but were lumped (along with A. semitorquatus) into A. taciturnus by Hellmayr (1938). Although part of the species complex has been examined in the past decades (Raposo 1997, Raposo & Parrini 1997), two subspecies (axillaris and nigrirostris) were not examined and neither were the vocalizations of most of these species, which could be a valuable tool for understanding the variations within the species complex.
In order to reevaluate the taxonomy, species limits, distribution and geographical variations in this species complex, we examined coloration and morphometrics of 1,041 specimens and 131 vocal recordings.
Our results suggest that there are 4 species involved in the complex: A. taciturnus, A. semitorquatus, A. franciscanus and A. axillaris, while A. t. nigrirostris is only part of a phenotypic variation found in A. taciturnus. These species have unique coloration patterns, mainly related to how much and which parts of the bill are yellow and/or black, the color of the anterior portions of the wings and the extent of superciliary stripes and pectoral band (Fig. 1, 2). They also have unique songs that can be used to distinguish them, except for A. axillaris, which overlaps with A. taciturnus in one song type. Despite having clear coloration and vocal differences, morphometrics are not sufficient to distinguishing the species, although some statistical differences in size can be observed.
 Figure 2 Ventral view of specimens: A. taciturnus (A [F, female], B [Male, M]); A. semitorquatus (C [M], D [F]), A. axillaris (E [F], F [M]); geographical variation of A. taciturnus with incomplete pectoral band (G [F], H [M]); A. franciscanus (I [M]). Note the buff coloration of the underparts and dusky pectoral band in female specimens
Figure 2 Ventral view of specimens: A. taciturnus (A [F, female], B [Male, M]); A. semitorquatus (C [M], D [F]), A. axillaris (E [F], F [M]); geographical variation of A. taciturnus with incomplete pectoral band (G [F], H [M]); A. franciscanus (I [M]). Note the buff coloration of the underparts and dusky pectoral band in female specimens
These birds are mostly endemic to particular regions with no or little overlap in their distributions (Fig. 1). Thus, A. semitorquatus is endemic to the Atlantic forests of southeastern Brazil; A. franciscanus is endemic to the threatened Caatinga in Northeastern Brazil; and A. axillaris is endemic to the east base of the northern Andes in Colombia and Venezuela; while A. taciturnus have a wider distribution occurring in the Amazon, the Atlantic forest and gallery forests with Amazonian influence in the Cerrado. How and when these species occupied such distinct environments is a question that is yet to be investigated. As a widespread species complex, further studies may help to clarify historical relationships between different biomes, which is a key to a broader and central question in evolutionary biology: Why are there so many species in the tropics?
In A. taciturnus, we have found complex intra-specific geographical variations in morphology and vocalization, which are partially incongruent. Populations from the Atlantic forest are very similar in plumage to populations from the Guyana Shields, having a complete pectoral band, while populations from the base of Andes have an incomplete pectoral band. All other populations have a great variety of intermediates, but also the pure phenotypes (Fig. 3). This suggests that, despite the pure phenotype from the extremes of distribution, gene flow may persist between the various populations. However, these variations are possibly a product of secondary contact or introgression between populations that have been historically separated.
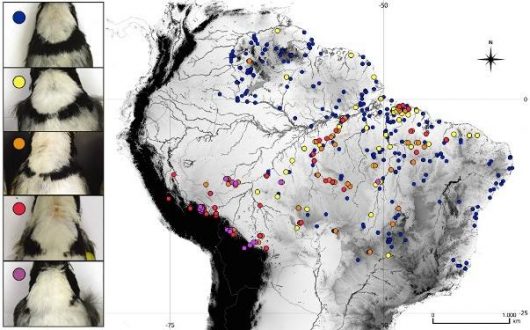 Figure 3 Geographic distribution of A. taciturnus specimens analyzed showing the highly polymorphic condition of the pectoral band extension from the complete phenotype (blue) to totally incomplete (purple). Note the tendency of populations from the base of the Andes, Inambari and Rondonia to have incomplete pectoral bands, whilst those from the Atlantic Forest and Guyana tend to have complete bands. All remaining populations are rich in intermediates
Figure 3 Geographic distribution of A. taciturnus specimens analyzed showing the highly polymorphic condition of the pectoral band extension from the complete phenotype (blue) to totally incomplete (purple). Note the tendency of populations from the base of the Andes, Inambari and Rondonia to have incomplete pectoral bands, whilst those from the Atlantic Forest and Guyana tend to have complete bands. All remaining populations are rich in intermediates
Regarding vocalizations, all populations pre-defined by areas of endemism (areas known for holding a high number of distinct taxa), were highly differentiable by a multivariate analysis. Although this is expected for oscine birds because they have a strong learning component in their song, i.e. they learn how to perform from their neighbors, the pattern observed doesn’t strictly agree with that. These birds don’t always sing more similarly to their adjacent populations. This suggests an additional influence factor that can be either due to genetic or habitat similarities. Also, contrary to the plumage analysis, vocal analyses didn’t corroborate the phenotype similarities between the Guyana and Atlantic Forest populations, but points to the “distinction” between populations from the base of the Andes (also seen in plumage) and more strikingly in populations from Belém (Fig. 4).
 Figure 4 Canonical analysis done with the vocal parameters of the different populations of A. taciturnus. In general, there was a tendency to form groups; however, there was overlap between most populations. Abbreviations correspond to the populations named after the areas of endemism: Atlantic Forest (ATF), Base of the Andes (BAN), Belém (BEL), Corridor of Dry Vegetation (CDV), Guy (Guyana Shields), Imeri (IME), Tapajós (TAP), Xingú (XIN)
Figure 4 Canonical analysis done with the vocal parameters of the different populations of A. taciturnus. In general, there was a tendency to form groups; however, there was overlap between most populations. Abbreviations correspond to the populations named after the areas of endemism: Atlantic Forest (ATF), Base of the Andes (BAN), Belém (BEL), Corridor of Dry Vegetation (CDV), Guy (Guyana Shields), Imeri (IME), Tapajós (TAP), Xingú (XIN)
Our study shows that just like many other cases in the Neotropics, the diversity in the Arremon genus is underestimated and there is still a lot to investigate concerning morphological and vocal variations in Neotropical birds. Although we expose some complex intra-specific variation patterns, further studies with molecular data are desirable and would be helpful to understand the (likely complex) evolutionary processes that drove the incongruent plumage and vocal patterns found in the different populations. This is being developed as part of my ongoing PhD, and hopefully will soon also shed light to some other hot questions such as how are the Atlantic forest, the Cerrado and the different areas in the Amazon are historically related in the context of biodiversity evolution.
References and further reading
Barrowclough, G.F., Cracraft, J., Klicka, J. & Zink, R.M. 2016. How many kinds of Birds are there and why does it matter? PLoS ONE 11(11): e0166307. VIEW
Hellmayr, C.E. 1938. Catalogue of birds of the Americas and adjacent islands. Field Museum Natural History Zoological Series no. 430, vol 13, part 11. Field Museum of Natural History, Chicago. VIEW
Raposo, M.A. 1997. A new species of Arremon (Passeriformes: Emberizidae) from Brazil. Ararajuba 5: 3–9. VIEW
Raposo, M.A. & Parrini, R. 1997. On the validity of the Half-collared Sparrow Arremon semitorquatus Swainson, 1827. Bulletin of the British Ornithologists’s Club 117: 294–298. VIEW
Image credit
Featured image: Pectoral Sparrow, Arremon taciturnus © Nelson Buainain
If you want to write about your research in #theBOUblog, then please see here.

