

LINKED PAPER
A pan-European, multipopulation assessment of migratory connectivity in a near-threatened migrant bird. Finch, T., Saunders, P., Avilés, J. M., Bermejo, A., Catry, I., de la Puente, J., Emmenegger, T., Mardega, I., Mayet, P., Parejo, D., Račinskis, E., Rodríguez-Ruiz, J., Sackl, P., Schwartz, T., Tiefenbach, M., Valera, F., Hewson, C., Franco, A. & Butler, S.J. 2015. Diversity and Distributions. DOI: 10.1111/ddi.12345
The loss of common and widespread bird species from the European countryside is becoming an increasingly familiar narrative (Inger et al. 2014). Two traits in particular seem to increase a species’ chances of having a negative conservation status; an association with farmed landscapes (Donald et al. 2006), and the habit of migrating to sub-Saharan African to see out the northern winter (Sanderson et al. 2006; Thaxter et al. 2010).
The European Roller Coracias garrulus is unfortunate enough to combine both of these traits, so its recent decline (Burfield & van Bommel, 2004; Birdlife International, 2015) should perhaps come as no surprise. The difficult part is figuring out which processes are to blame. Whilst most conservationists point to the intensification of Europe’s farmed landscape (Kovacs et al. 2008), we should also consider whether the Roller is affected by problems during the non-breeding season. This is where things might get complicated, with recent studies suggesting that migrant populations are influenced by factors interacting throughout the migratory cycle, the effects of which may vary within and between species (Morrison et al. 2013; Vickery et al. 2013).
Tracking Roller movements
The first step towards assessing non-breeding season threats is an understanding of the year-round movements of our Rollers. Until recently the only such information came from ring recoveries, which proved to be a frustratingly slow way of gathering Roller migration data; a Bulgarian bird found dead in Tanzania in 1936 remains our only sub-Saharan ring recovery. Mercifully, our reliance on ringing came to end when animal tracking technology – once reserved for only the largest of species – became sufficiently lightweight for use with smaller birds (Bridge et al. 2011).
 Roller recaptured with geolocator ready to be removed and downloaded © Simon Butler
Roller recaptured with geolocator ready to be removed and downloaded © Simon Butler
By collaborating with other Roller researchers, we’ve been able to combine our own Roller migration data (available at the Movebank Data Repository) with all existing data from solar geolocators and ‘PTT’ satellite tags (Catry et al. 2014; Emmenegger et al. 2014; Rodríguez-Ruiz et al. 2015), as well as the solitary sub-Saharan ring recovery. So, after waiting 80 years for a second winter ring recovery, we have now amassed data on the year-round movements of 31 Rollers from seven countries across Europe. Our open access paper tells the story of these heroic thirty-one.
Autumn migration
The key finding here is the stopover made by all birds in the northern savannah, between the Sahara desert and the equatorial rainforest. These stopovers lasted about a month (between mid-September and early November), representing a key refuelling period. Due to the spatial imprecision of geolocator data, we’re not exactly sure which sites the rRllers are using, but they seem to be concentrating towards the Lake Chad basin, which is certainly used by the Spanish birds tracked using satellite tags.
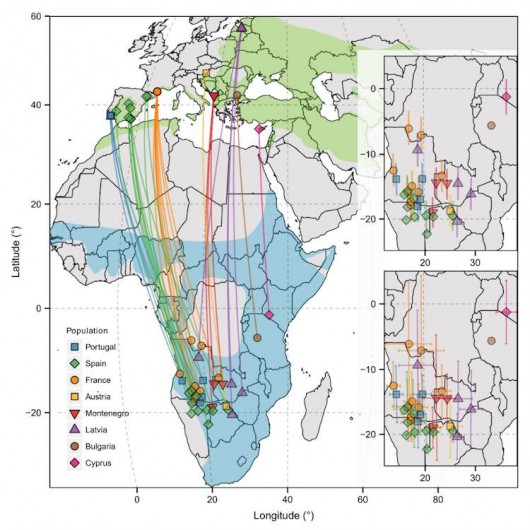 Straight lines (not actual routes) connect breeding and winter sites of tracked Rollers. Inset maps indicate uncertainty around the winter positions. See paper for more details.
Straight lines (not actual routes) connect breeding and winter sites of tracked Rollers. Inset maps indicate uncertainty around the winter positions. See paper for more details.
Winter
After this stopover, all Rollers continued across the equator to winter in the grasslands and shrublands of southern Africa. Unlike many European migrants which winter north of the equator during the dry season (e.g. in the Sahel), the rollers’ arrival in southern Africa coincides with the return of the rainy season (click here for a cool visualisation of the Inter-Tropical Convergence Zone, which drives this seasonality). This presumably makes their epic journey (4,000 – 8,000 km as the crow flies) worthwhile.
How did Rollers from across Europe distribute themselves in Africa? This is a key question, encompassed by the term ‘migratory connectivity’ (Webster et al. 2002). Briefly, connectivity is described as ‘strong’ when individuals from different breeding populations are segregated during the non-breeding season. This might mean that, say, Latvian Rollers all overwinter together in the same small part Africa, encountering similar conditions year-round and thus being vulnerable (at the population level) to any environmental change at this site. ‘Weak’ (or diffuse) connectivity, on the other hand, describes the opposite scenario whereby birds from different breeding populations mix up over winter. Latvian Rollers might spread out over a larger part of Africa, being more resilient (at the population level) to environmental change. They will also encounter birds from elsewhere in Europe, so perhaps their chances of surviving the winter will be affected by how many French Rollers are about to compete with. This is all hypothetical, but demonstrates the ways in which migratory connectivity might be important for understanding migrant population dynamics (Dolman & Sutherland, 1992; Norris & Marra, 2007; Jones et al. 2008).
In our case, migratory connectivity was somewhere in the middle of this weak-strong continuum. Whilst there was some mixing of Rollers from different breeding populations, their winter distribution wasn’t completely random. In other words, a Roller was likely to winter close to a Roller that it bred near to, and far from a Roller that it bred far from. This connectivity was driven by two patterns. The first, ‘parallel migration’, is apparent in the strong correlations between the longitude of breeding and winter sites; Rollers from western Europe wintered in western Africa, and those from eastern Europe wintered in eastern Africa. The second pattern, ‘leap-frog migration’ (which operated in the east, but not in the west), occurs when northern breeders winter to the south of the winter range. In our case, Latvian rollers leap-frogged over the Cypriot bird, wintering around 2,000 km further south.
Spring migration
All individuals showed, to some extent, a ‘loop migration’, with spring routes differing from autumn routes. This pattern was most striking for Latvian Rollers, whose northbound journey took them over the horn of Africa and along the Arabian Peninsula. Ring recoveries (remember them?) from Latvia, Hungary and Serbia suggest that this easterly spring route is not unique to our tagged Latvian Rollers. Anecdotal evidence indicates that hunting is major problem for spring migrants in Arabia, and tracking is telling us which breeding populations are exposed to these threats.
 Rollers shot on spring migration through Arabia in 2015. Photo from Twitter user @Khalidababatain (not the perpetrator!), via @JamieDunning
Rollers shot on spring migration through Arabia in 2015. Photo from Twitter user @Khalidababatain (not the perpetrator!), via @JamieDunning
Conclusion
I hope I’ve convinced you of the value of combining avian tracking data from across a species’ range. Beyond getting all the data on one map, this pan-European effort has allowed us to perform some quantitative analyses of migratory connectivity. Whilst we haven’t solved the mystery of the Roller’s decline, we’ve made a decent step towards understanding their non-breeding movements. We can now start asking how non-breeding conditions affect the demography of our study populations, and ultimately figure out whereabouts in the migratory cycle population size is being limited. More generally, our work complements other recent wide-scale tracking studies (e.g. Stanley et al. 2014; Ouwehand et al. 2015), allowing us to question the broader causes and consequences of variation in migration strategy.
References and further reading
Birdlife International (2015) European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. View
Bridge, E.S., Thorup, K., Bowlin, M.S., Chilson, P.B., Diehl, R.H., Fléron, R.W., Hartl, P., Kays, R., Kelly, J.F., Robinson, W.G. & Wikelski, M. (2011) Technology on the Move: Recent and Forthcoming Innovations for Tracking Migratory Birds. BioScience 61: 689–698. View
Burfield I. & van Bommel F. (2004) Birds in Europe: Population Estimates, Trends and Conservation Status. BirdLife International, Cambridge, UK.
Catry I., Catry T., Granadeiro J.P., Franco A.M.A., & Moreira F. (2014) Unravelling migration routes and wintering grounds of European rollers using light-level geolocators. Journal of Ornithology 155: 1071–75. View
Dolman P.M. & Sutherland W.J. (1992) The response of bird populations to habitat loss. Ibis 137: S38–S46. View
Donald P.F., Sanderson F.J., Burfield I.J., & van Bommel F.P.J. (2006) Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agriculture, Ecosystems & Environment 116: 189–196. View
Emmenegger T., Mayet P., Duriez O., & Hahn S. (2014) Directional shifts in migration pattern of rollers (Coracias garrulus) from a western European population. Journal of Ornithology 155: 427–433. View
Inger R., Gregory R., Duffy J.P., Stott I., Voříšek P., & Gaston K.J. (2014) Common European birds are declining rapidly while less abundant species’ numbers are rising. Ecology Letters 18: 28–36. View
Jones J., Norris D.R., Girvan M.K., Barg J.J., Kyser T.K., & Robertson R.J. (2008) Migratory connectivity and rate of population decline in a vulnerable songbird. The Condor 110: 538–544. View
Kovacs A., Barov B., Orhun C., & Gallo-Orsi U. (2008) International Species Action Plan for the European Roller Coracius garrulus garrulus. Besenyőtelek, Hungary. View
Morrison C.A., Robinson R.A., Clark J.A., Risely K., Gill J.A., & Duncan R. (2013) Recent population declines in Afro-Palaearctic migratory birds: the influence of breeding and non-breeding seasons. Diversity and Distributions 19: 1051–1058. View
Norris D.R. & Marra P.P. (2007) Seasonal interactions, habitat quality, and population dynamics in migratory birds. The Condor 109: 535–547. View
Ouwehand J., Ahola M.P., Ausems A.N.M.A., Bridge E.S., Burgess M., Hahn S., Hewson C., Klaassen R.H.G., Laaksonen T., Lampe H.M., Velmala W., & Both C. (2015) Light-level geolocators reveal migratory connectivity in European populations of pied flycatchers Ficedula hypoleuca. Journal of Avian Biology. DOI: 10.1111/jav.00721. View
Rodríguez-Ruiz J., de la Puente J., Parejo D., Calero-Torralbo M.Á., Reyes-González J.M., Zajková Z., Bermejo A., & Avilés J.M. (2015) Disentangling migratory routes and wintering grounds of Iberian near-threatened European Rollers Coracias garrulus. PLoS ONE. DOI: 10.1371/journal.pone.0115615. View
Sanderson F.J., Donald P.F., Pain D.J., Burfield I., & van Bommel F. (2006) Long-term population declines in Afro-Palearctic migrant birds. Biological Conservation 131: 93–105. View
Stanley C.Q., McKinnon E.A., Fraser K.C., Macpherson M.P., Casbourn G., Friesen L., Marra P.P., Studds C., Ryder T.B., Diggs N.E., & Stutchbury B.J.M. (2014) Connectivity of wood thrush breeding, wintering, and migration sites based on range-wide tracking. Conservation Biology 29: 164–174. View
Thaxter C.B., Joys A.C., Gregory R.D., Baillie S.R., & Noble D.G. (2010) Hypotheses to explain patterns of population change among breeding bird species in England. Biological Conservation 143: 2006–2019. View
Vickery J.A., Ewing S.R., Smith K.W., Pain D.J., Bairlein F., Škorpilová J., & Gregory R.D. (2013) The decline of Afro-Palaearctic migrants and an assessment of potential causes. Ibis 156: 1–22. View
Webster M.S., Marra P.P., Haig S.M., Bensch S., & Holmes R.T. (2002) Links between worlds: unravelling migratory connectivity. TRENDS in Ecology & Evolution 17: 76–83. View
Image credits
European Roller in the hand © Tom Finch; Tom Finch © Simon Butler
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here.

