

 LINKED PAPER
LINKED PAPER
Evaluating the ability of occurrence models to predict nest locations and associated vegetation. Fogarty, F.A., Fleishman, E. & Zillig, M.W. 2021 IBIS. doi: 10.1111/ibi.13020 VIEW
The data required to estimate reproductive status or success, especially for relatively rare species, can be expensive, invasive and difficult to obtain. Occurrence data, which do not rely on capturing individual birds, are frequently used as an easier alternative to infer characteristics of habitat and inform management plans for large areas. However, the extent to which these data predict reproduction, or the environmental characteristics association with reproduction, is uncertain. Given that occurrence data are more widely available and much less labor-intensive to collect than data on nest locations or reproductive metrics, and that many management plans are based on general associations between bird species and coarse-resolution land-cover or vegetation types, there is practical relevance in evaluating whether inferences about nesting can be derived from occurrence models.
Although it often is assumed that the occurrence of a species at a given location during the breeding season is an indication of breeding activity, there are several reasons why detection at a particular site may not imply breeding or reproductive success at the same site. For example:
- A survey site may partially overlap with one or more territories of a species, but the overlapping portion may not meet the species’ nesting requirements (e.g., the nest may be located outside the site).
- A species’ abundance in high-quality breeding habitat may be relatively low if highly competitive individuals establish territories and exclude other individuals (Van Horne 1983).
- A site may be occupied by non-breeding individuals. These could be birds that did not pair or breed, or individuals of species that have multiple broods and are searching for a new nest location.
- Unpaired males may occupy lower quality habitat and be more detectable than paired males because they sing more frequently (Gibbs and Wenny 1993).
- Abundance may be negatively correlated with reproductive output by mechanisms such as ecological traps, wherein rapid environmental change produces uninformative cues that induce birds to select lower-quality habitat (Pidgeon et al. 2003; Remeš 2003).
We tested several hypotheses to examine whether models built from occupancy data were reasonable surrogates for models built from data on specific nest locations. We used data on occurrence and nest locations for three species (Fig. 1) in the montane western Great Basin, USA (Fig. 2) which we classified as either nest-stratum generalists or specialists. We chose two nest-stratum generalists: Brewer’s Sparrows (Spizella breweri), which nest in extensive, nearly treeless shrub steppe and use a variety of abundant woody shrubs as nest strata; and Green-tailed Towhees (Pipilo chlorurus), which nest in similar shrubs or on the ground and peak in abundance in ecotones between shrub steppe and woodlands. We classified American Dusky Flycatchers (Empidonax oberholseri) as nest-stratum specialists in this region given that they nest in forest openings, riparian vegetation, and patches of small deciduous trees, which are relatively scarce, and typically construct nests in small, deciduous trees.

Figure 1 Top: Barrett Canyon in the Shoshone Mountains of central Nevada, USA. The extensive shrub steppe pictured is used as nesting habitat for Brewer’s Sparrow (Spizella breweri) and Green-tailed Towhee (Pipilo chlorurus), while American Dusky Flycatcher (Empidonax oberholseri) nest in the scattered Cercocarpus ledifolius woodland higher up the slope. Bottom: Our three focal species (from left to right) Green-tailed Towhee, American Dusky Flycatcher and Brewer’s Sparrow.
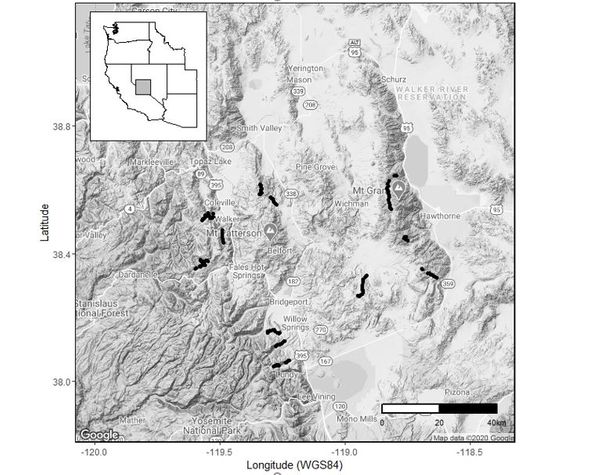
Figure 2 Point count sites in the western Great Basin. Black circles represent 154 locations in 13 canyons in the Sierra Nevada (7 canyons), Sweetwater Mountains (2), Bodie Hills (1), and Wassuk Range (3).
First, we compared whether vegetation structure and composition within 50 m of occurrence and nest locations for each species were similar. We expected that the vegetation attributes associated with occurrence and nest location of our nest-stratum generalists would be more similar than those of the nest-stratum specialist because the former are less reliant on nest strata that are scarce. Second, we used data on the vegetation within 50 m of known nests to evaluate whether occurrence models accurately predicted whether nests were present in a given location. We also anticipated that occurrence models would classify the presence of the nests of the nest-stratum generalists more accurately than those of the nest-stratum specialist. Third, we examined whether nest strata (vegetation composition and structure within 5 m of a known nest) differed from that at randomly chosen locations of the same size within 20–80 m of the nest. We predicted that the nest strata of American Dusky Flycatchers would be more distinct from the surrounding area than those of Brewer’s Sparrows and Green-tailed Towhees, indicating that the former are using specific nest resources that are less abundant.
We found that models based on occurrence data suggested different vegetation variables were important when compared to models based on nest site data for all three species. However, our occurrence models classified nest locations with relatively high accuracy for Brewer’s Sparrow (AUC = 0.96) and American Dusky Flycatcher (AUC = 0.93), and with moderate accuracy for Green-tailed Towhee (AUC = 0.70), indicating that these occurrence models effectively indicated whether the species were nesting within a given 100 m radius (Fig 2). Contrary to our hypotheses, the occurrence model for our nest stratum specialist (American Dusky Flycatcher) performed the best overall at predicting nesting (accuracy, sensitivity, and specificity all > 0.84). Vegetation within 5 m of nests also differed from that of random locations within 20–80 m for all three species, suggesting that even the nest stratum generalists based nest selection on fine-grained attributes.
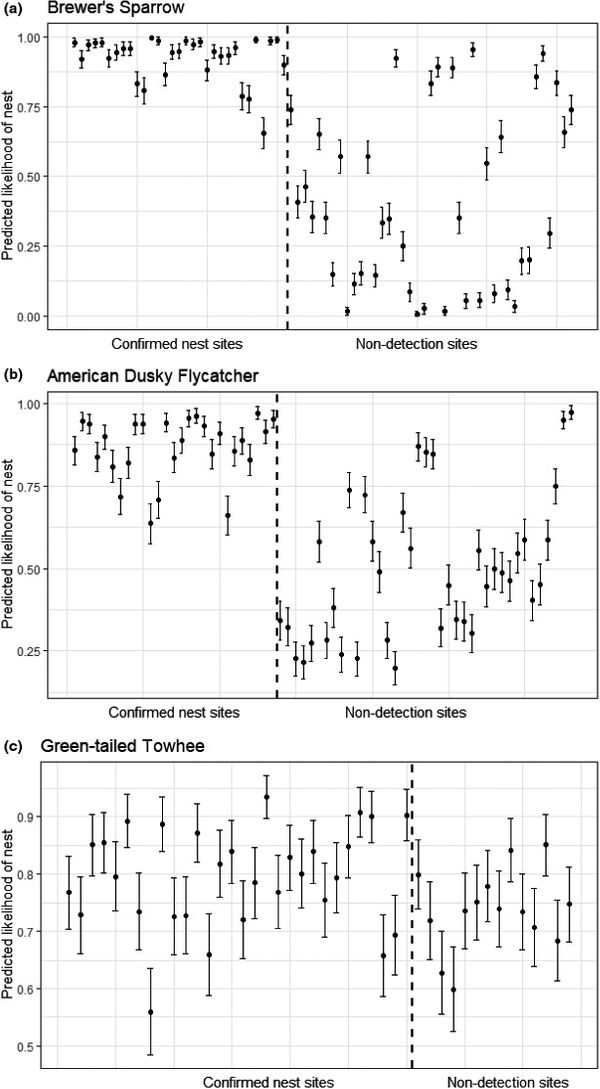
Figure 3 Predicted likelihood of nesting of (A) Brewer’s Sparrow, (B) American Dusky Flycatcher, and (C) Green-tailed Towhee. Predictions were generated from models built with occurrence data and then fit to vegetation data from nest location data. Black circles represent the mean predicted likelihood for a given point location and vertical bars represent the standard error of that estimate. Points to the left of the vertical dashed line are sites with nests and points to the right of the vertical dashed line are non-detection sites.
Although there were substantial differences in attributes of vegetation composition and structure associated with occurrence and nesting locations, our models of occurrence effectively classified the presence or absence of nesting with high accuracy for two of our three species, suggesting that models associating occurrence data with environmental variables may be useful for inferring the presence or quality of nesting habitat for these species in lieu of more labor-intensive nest searching.
However, these occurrence models were fit to relatively fine-resolution, site-specific vegetation data. Derivation of valid inferences about nesting habitat is predicated on a well-fitting model that accurately predicts occurrence at a meaningful spatial scale. We suggest that assumptions about the correlation between point-count data and nesting be validated for given study systems before inferring the quality of nesting habitat on the basis of point count data. Identifying additional species for which occurrence models can predict nest presence could be valuable for environmental compliance, multiple-use management, and songbird conservation efforts.
References
Gibbs, J.P. & Wenny, D.G. 1993. Song output as a population estimator: effect of male pairing status. Journal of Field Ornithology 64: 316-322. VIEW
Pidgeon, A., Radeloff, V. & Mathews, N. 2003. Landscape-scale patterns of Black-throated Sparrow (Amphispiza bilineata) abundance and nest success. Ecological Applications 13: 530-542. VIEW
Remeš, V. 2003. Effects of exotic habitat on nesting success, territory density, and settlement patterns in the Blackcap (Sylvia atricapilla). Conservation Biology 17: 1127-1133. VIEW
Van Horne, B. 1983. Density as a misleading indicator of habitat quality. Journal of Wildlife Management 47: 893-901. VIEW
Image credit
Top right: Green-tailed Towhee Pipilo chlorurus © Frank Fogarty.
If you want to write about your research in #theBOUblog, then please see here.

