

LINKED PAPER
Misidentifications in citizen science bias the phenological estimates of two hard-to-identify Elaenia flycatchers. Gorleri, F.C. & Areta, J.I. 2021 IBIS. DOI: 10.1111/ibi.12985 VIEW
 Citizen science databases are increasingly used to answer a variety of ecological questions for birds, but we still have a poor understanding of how identification errors may affect the resulting output of analyses. The potential for species misidentifications between hard-to-identify taxa is certainly one limitation of citizen science databases, as non-expert observers who largely participate as data-collectors in these initiatives might be more prone to misidentify birds. Of particular concern is that, if identification errors are produced systematically and at broad scales, the noisy data may compromise ecological estimates derived from them. Surprisingly, however, the identification accuracy of databases remains a largely unexplored topic, and thus, the effects of misidentified reports during data analysis remain poorly understood.
Citizen science databases are increasingly used to answer a variety of ecological questions for birds, but we still have a poor understanding of how identification errors may affect the resulting output of analyses. The potential for species misidentifications between hard-to-identify taxa is certainly one limitation of citizen science databases, as non-expert observers who largely participate as data-collectors in these initiatives might be more prone to misidentify birds. Of particular concern is that, if identification errors are produced systematically and at broad scales, the noisy data may compromise ecological estimates derived from them. Surprisingly, however, the identification accuracy of databases remains a largely unexplored topic, and thus, the effects of misidentified reports during data analysis remain poorly understood.
Flycatchers (family Tyrannidae) are known to cause identification headaches to birdwatchers and scientists alike (Pearman and Areta, 2021). In the Neotropical region, several species of Elaenia flycatchers that are extremely similar in appearance co-exist, and species-level ID is challenging even for experienced observers. Two species, the taxonomically debated Chilean Elaenia (Elaenia chilensis) and the Small-billed Elaenia (E. parvirostris) are migratory and partially overlap in space and time (see Figure 1); therefore, reciprocal identification errors likely occur when they are found together. Although each species breeds in separate but sometimes adjacent regions in southern South America, they largely overlap ranges during their migratory periods, when Chilean Elaenia migrates through the breeding range of Small-billed Elaenia during autumn and spring migration (Figure 1). Given this distributional particularity, we were interested in evaluating the identification accuracy of their photographic reports in citizen science databases, and test whether misidentifications produce any shift in their estimates of migration phenologies across the region where their ranges overlap.
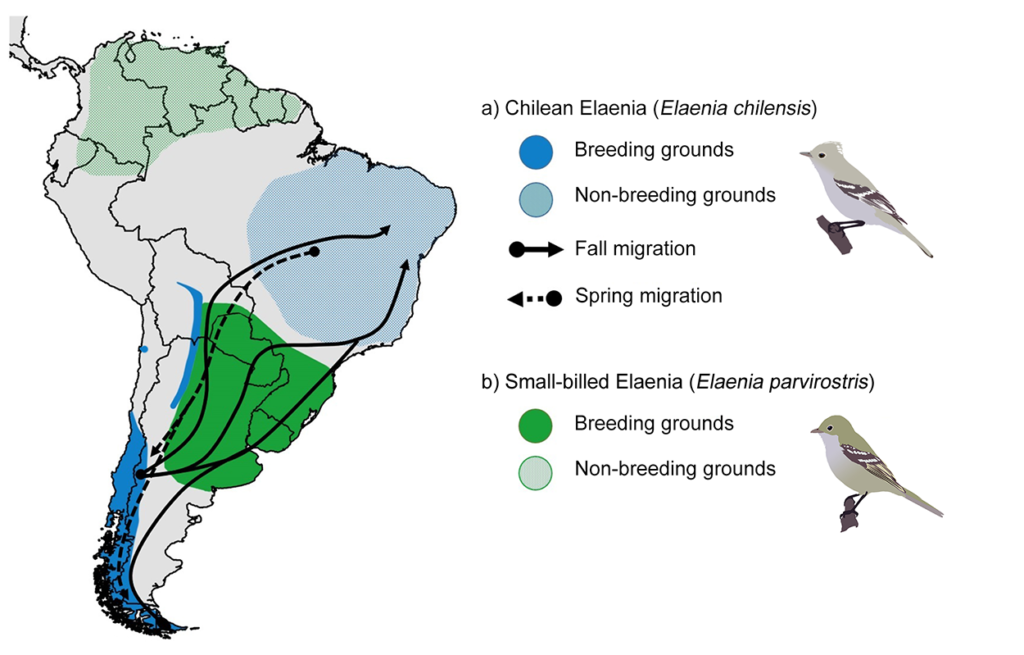 Figure 1 Schematic distribution maps of Chilean Elaenia Elaenia chilensis and Small-billed Elaenia E. parvirostris, and migration routes of Chilean Elaenia. Migration routes are based on geolocator data given in Jiménez et al. (2016) and Bravo et al. (2017). Note that Chilean Elaenia migrates over the breeding range of Small-billed Elaenia.
Figure 1 Schematic distribution maps of Chilean Elaenia Elaenia chilensis and Small-billed Elaenia E. parvirostris, and migration routes of Chilean Elaenia. Migration routes are based on geolocator data given in Jiménez et al. (2016) and Bravo et al. (2017). Note that Chilean Elaenia migrates over the breeding range of Small-billed Elaenia.
How many misidentified photos are there?
We reviewed 4399 photos of both Chilean Elaenia and Small-billed Elaenia submitted to eBird and EcoRegistros in Chile, Argentina and Uruguay in search of misidentified reports, and explored how errors were distributed in space and time. We found few wrong reports (27 of 1395, 2%) in photos that were uploaded as Chilean Elaenia, suggesting a high data accuracy in the photographic reports in this species. By contrast, we found a large number of errors (173 of 1348, 11.4%) in photos that were uploaded as Small-billed Elaenia, 113 of which actually belonged to Chilean Elaenia.
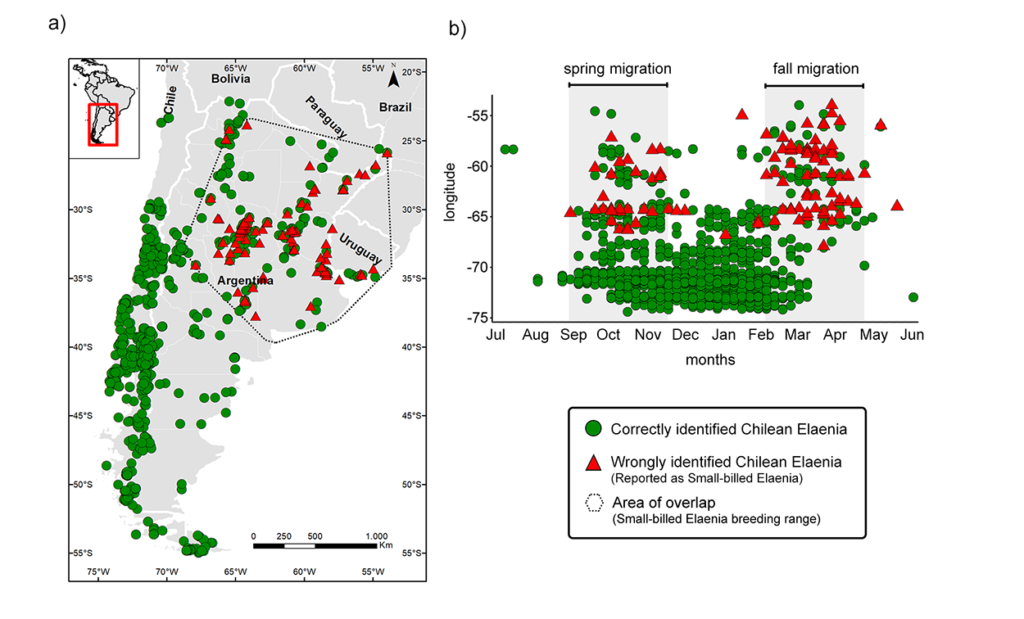 Figure 2 Spatio-temporal distribution of correctly identified Chilean Elaenia Elaenia chilensis (green circles) and misidentified Chilean Elaenia reported as Small-billed Elaenia E. parvirostris (red triangles) in eBird and EcoRegistros for Argentina, Chile and Uruguay. (a) Geographical distribution of reports and (b) temporal distribution of reports as a function of longitude. Note that errors concentrate during the spring and autumn (fall) migratory periods when both species overlap in the breeding range of Small-billed Elaenia.
Figure 2 Spatio-temporal distribution of correctly identified Chilean Elaenia Elaenia chilensis (green circles) and misidentified Chilean Elaenia reported as Small-billed Elaenia E. parvirostris (red triangles) in eBird and EcoRegistros for Argentina, Chile and Uruguay. (a) Geographical distribution of reports and (b) temporal distribution of reports as a function of longitude. Note that errors concentrate during the spring and autumn (fall) migratory periods when both species overlap in the breeding range of Small-billed Elaenia.
The errors were not evenly distributed through space and time, rather they concentrated on the overlapping area of Chilean and Small-billed Elaenias and at times of geographic overlap during the migratory periods (Figure 2). Interestingly, we found that erroneous reports of Small-billed Elaenia that belonged to Chilean Elaenia were mostly concentrated in autumn rather than spring (Figure 2). This skewed distribution of errors towards the autumn migration period resulted from confusion with juveniles of Chilean Elaenia, that are particularly abundant during the post-breeding (autumn) migration and are harder to identify than adults.
Do errors affect the migration phenology estimates in these flycatchers?
Yes they do! The large number of wrong reports that concentrate toward the autumn migration period produced shifts in the estimates that we made about the migration phenology in these species. These shifts were clearly visible when we compared a phenological model built using the raw data (data containing all errors) to a model built with fully curated data (identification error-free data; Figure 3).
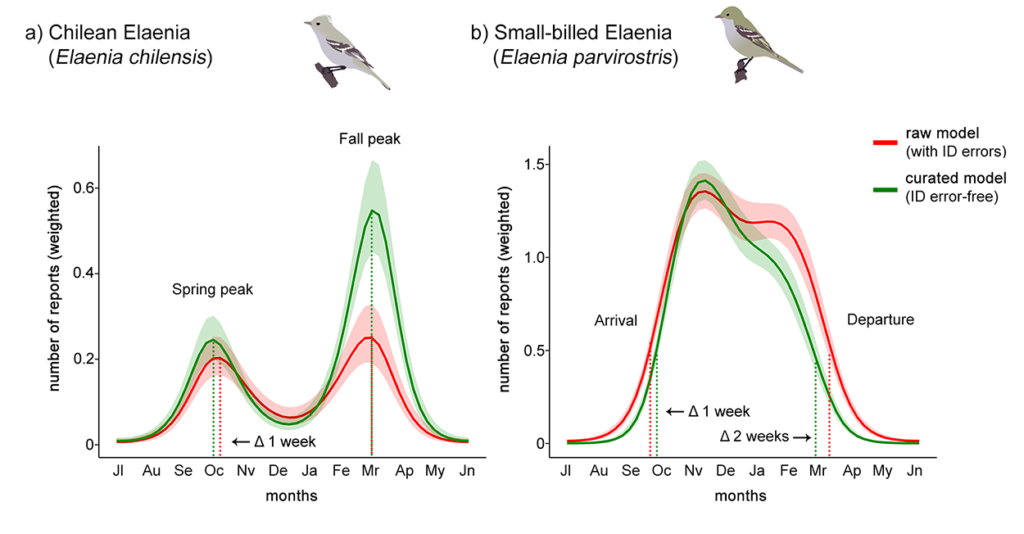 Figure 3 Phenological GAMs of (a) Chilean Elaenia Elaenia chilensis and (b) Small-billed Elaenia E. parvirostris built with raw citizen science reports (red lines) and curated citizen science reports (green lines), across the overlapping area of both species in Argentina and Uruguay (see Fig. 1). Dotted lines indicate the estimated peak of spring and autumn (fall) migration for Chilean Elaenia, and estimated arrival/departure dates for Small-billed Elaenia. The shaded area represents the 95% confidence interval of each model. Curated models estimated a 1-week difference in the spring peak passage and a stronger autumn peak in Chilean Elaenia, and a 1-week later arrival and a 2-week earlier departure in Small-billed Elaenia.
Figure 3 Phenological GAMs of (a) Chilean Elaenia Elaenia chilensis and (b) Small-billed Elaenia E. parvirostris built with raw citizen science reports (red lines) and curated citizen science reports (green lines), across the overlapping area of both species in Argentina and Uruguay (see Fig. 1). Dotted lines indicate the estimated peak of spring and autumn (fall) migration for Chilean Elaenia, and estimated arrival/departure dates for Small-billed Elaenia. The shaded area represents the 95% confidence interval of each model. Curated models estimated a 1-week difference in the spring peak passage and a stronger autumn peak in Chilean Elaenia, and a 1-week later arrival and a 2-week earlier departure in Small-billed Elaenia.
For Small-billed Elaenia, errors caused a 1-week advancement in the estimated arrival (late September instead of early October), and a 2-week delay in the estimated departure (early April instead of mid-March). For Chilean Elaenia, errors caused a 1-week delay for the estimated spring peak passage (mid-October instead of early October), and an underestimation of the magnitude of the autumn passage that occurred mostly from March to April. We expected this outcome, as most of the wrong reports of Small-billed Elaenia that belonged to Chilean were concentrated from March to May. This uneven distribution of errors overestimated the presence of Small-billed Elaenia towards the end of the migratory period, and at the same time, underestimated the autumn migratory passage of Chilean Elaenia.
Take-home message
We commend the growing ability of citizen science databases to describe spatio-temporal patterns in birds. However, we also advertise about the need to undertake active post-processing of records before analysing data, especially so when the species of interest is hard to identify (see also Areta & Juhant 2019). Large-scale analyses involving many taxa often do not allow for manual correction or testing of identification accuracy. Therefore, we believe that citizen science initiatives could do well in alerting users and researchers to the most commonly misidentified species or species groups in their databases. While scientists are ultimately responsible for the data used for analysis, this warning on potential problems arising from species misidentifications can help mitigate the misuse of raw data in phenological or other analyses, thereby increasing the value of citizen science data.
Image credit
Top right: Chilean Elaenia Elaenia chilensis © Pablo Eguia
References
Areta, J.I. & Juhant, M.A. 2019. The Rufous-thighed Kite Harpagus diodon is not an endemic breeder of the Atlantic Forest: lessons to assess Wallacean shortfalls. Ibis 161: 337– 345. VIEW
Bravo, S.P., Cueto, V.R. & Gorosito, C.A. 2017. Migratory timing, rate, routes and wintering areas of Chilean Elaenia (Elaenia albiceps chilensis), a key seed disperser for Patagonian Forest regeneration. PLoS One 12: 1–15. VIEW
Jiménez, J.E., Jahn, A.E., Rozzi, R. & Seavy, N.E. 2016. First documented migration of individual Chilean Elaenias (Elaenia albiceps chilensis) in South America. Wilson Journal of Ornithology 128: 419–425. VIEW
Pearman, M. & Areta, J.I. 2021. Field identification of some look-alike Serpophaga tyrannulets and Plain Inezia from Argentina. Neotropical Birding 28: 28–33. VIEW

