

LINKED PAPER
Elevational plumage divergence in the Rufous-capped Babbler (Cyanoderma ruficeps) on a mountainous island. Fang, Y-T., Yao, C-T., Hsu, Y-C. & Hung, C-M. 2021 IBIS. doi: 10.1111/ibi.13009 VIEW

During the summer in Taiwan, people enjoy driving up to high mountains for a moment of coolness because the lowland is so hot. On this subtropical, mountainous island, it takes only a few hours to drive from sea-level to 3000-metre-high mountains. Along the trip, you may find habitats changing from subtropic lowland forests to coniferous forests, and then mountain grasslands (Fig. 1). However, it is still possible to get a suntan or even become sunburnt in the cool, high mountains. This is because the levels of UV radiation from the sun are high at these altitudes and often higher than those in the lowlands. It is unclear how species, including birds, adapt evolutionarily to this change in the light environment observed between lowland and mountainous landscapes.
 Figure 1 Characteristic lowland (A) and high-altitude mountainous habitats (B) of Rufous-capped Babblers (Cyanoderma ruficeps) in Taiwan. This species often occurs in tall grasses or bushes of various types across many habitats, such as (A) subtropical forest and (B) coniferous woodland.
Figure 1 Characteristic lowland (A) and high-altitude mountainous habitats (B) of Rufous-capped Babblers (Cyanoderma ruficeps) in Taiwan. This species often occurs in tall grasses or bushes of various types across many habitats, such as (A) subtropical forest and (B) coniferous woodland.
The plumage colouration of birds may have evolved for intraspecific communication but may also have been constrained by predation risk. Given that plumage signal transmission is mediated by the background environment, different habitats may cause plumage colouration to diverge between populations or species (Bertrand et al. 2016; Medina et al. 2017). Previous studies have shown environmental impacts on plumage colourations at various scales, such as forest layers and latitudinal zones, among species (Gomez & Théry 2004; Johnson et al. 2013). However, few studies have examined plumage evolution across an elevational gradient, along which ecological conditions such as UV light environments may change. One interesting question is: do differences in ambient UV light lead to plumage UV reflectance differences between birds from lowland and montane habitats?
In 2017, we aimed to answer this question by examining whether a widespread passerine, the Rufous-capped Babbler (Cyanoderma ruficeps), showed changes in plumage colouration across elevations in Taiwan. This bird has pale olive or dark grey plumage covering most of its body parts, but also has a red cap, a yellow throat and belly, and white or yellow lesser coverts. We cannot tell any plumage differences between lowland and montane individuals or between males and females by-eye. Thus, Yi-Ting Fang, who is the first author of this study and was then an undergraduate student, used a spectrometer to measure the plumage colouration of specimens, collected from lowland and mountain habitats (Fig. 1), archived at the Endemic Species Research Institute in Taiwan. She analysed the reflectance spectral data based on two systems—objective colorimetry (Hill & McGraw 2006) and tetrahedral colour space (Stoddard & Prum 2008).
To assess differences in UV light environments across elevations in Taiwan, we obtained ultraviolet index (UVI) data from various locations collected by the Central Weather Bureau of Taiwan. Combining the plumage reflectance and UVI data, we showed that Rufous-capped Babblers had higher UV-reflecting and brighter plumage in mountainous habitats, where ambient UV light is stronger, than lowland habitats (Fig. 3 & 4). The elevational differences were found in their ventral parts, which already had relatively high levels of UV reflectance but not in their dorsal parts, which were likely subject to high aerial predation risk. The results imply that the plumage evolution of Rufous-capped Babblers is regulated by both intraspecific communication and predation risk.
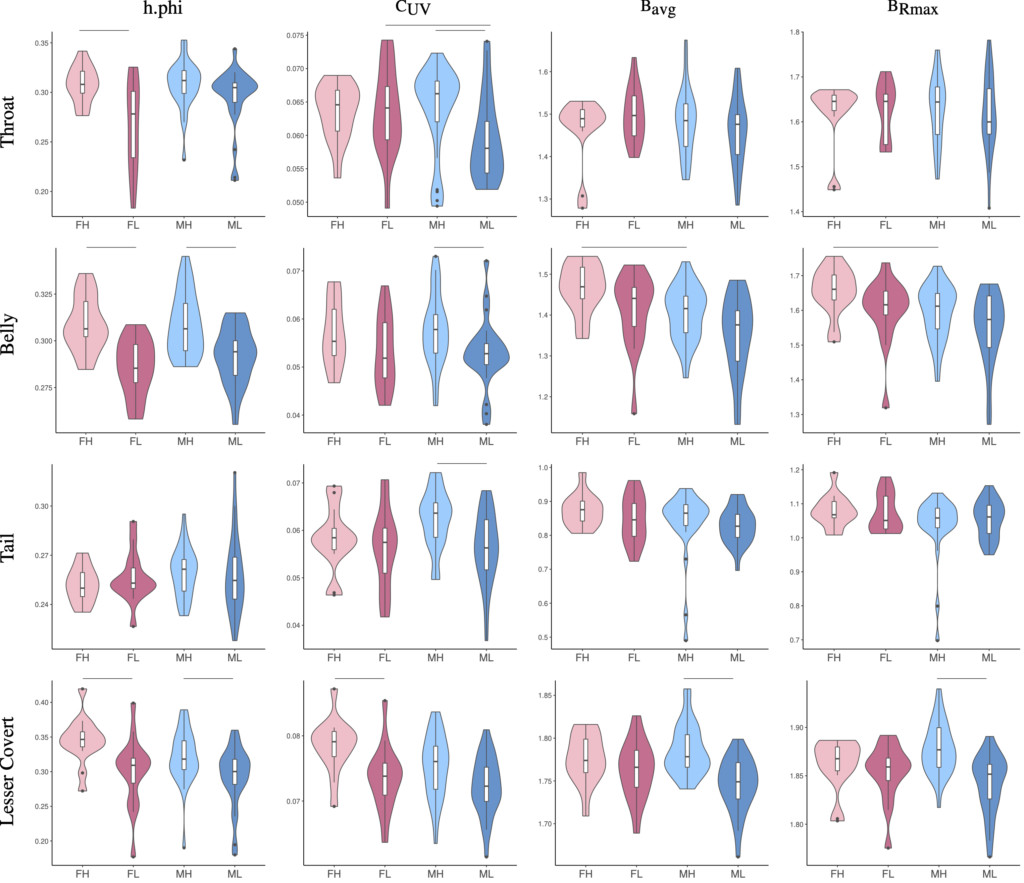 Figure 2 Comparisons of UV or brightness-associated plumage reflectance parameters — h.phi, CUV, BRmax, and Bavg — between the sexes and across elevations for the bellies and lesser coverts of Rufous-capped Babblers. Light and dark pink colours indicate females from mountainous (FH) and lowland habitats (FL), respectively. Light and dark blue colours indicate males from mountainous (MH) and lowland habitats (ML), respectively. Horizontal bars indicate significant Mann-Whitney U-test results with Benjamini-Hochberg adjusted P-values < 0.05.
Figure 2 Comparisons of UV or brightness-associated plumage reflectance parameters — h.phi, CUV, BRmax, and Bavg — between the sexes and across elevations for the bellies and lesser coverts of Rufous-capped Babblers. Light and dark pink colours indicate females from mountainous (FH) and lowland habitats (FL), respectively. Light and dark blue colours indicate males from mountainous (MH) and lowland habitats (ML), respectively. Horizontal bars indicate significant Mann-Whitney U-test results with Benjamini-Hochberg adjusted P-values < 0.05.
We also found sexual dichromatism in different body parts between montane and lowland babblers. We believe that the different patterns of sexual dichromatism across elevation is more likely caused by unequal natural selection pressures between sexes than by sexual selection. This is because we found that females tended to have brighter and stronger UV-associated colouration than males (Fig. 3), which conflicts with the pattern predicted by sexual selection. Given that females usually prefer more colourful or brighter males that may be healthier, sexual selection by female choice should lead to brighter males than females (Dunn et al. 2015).
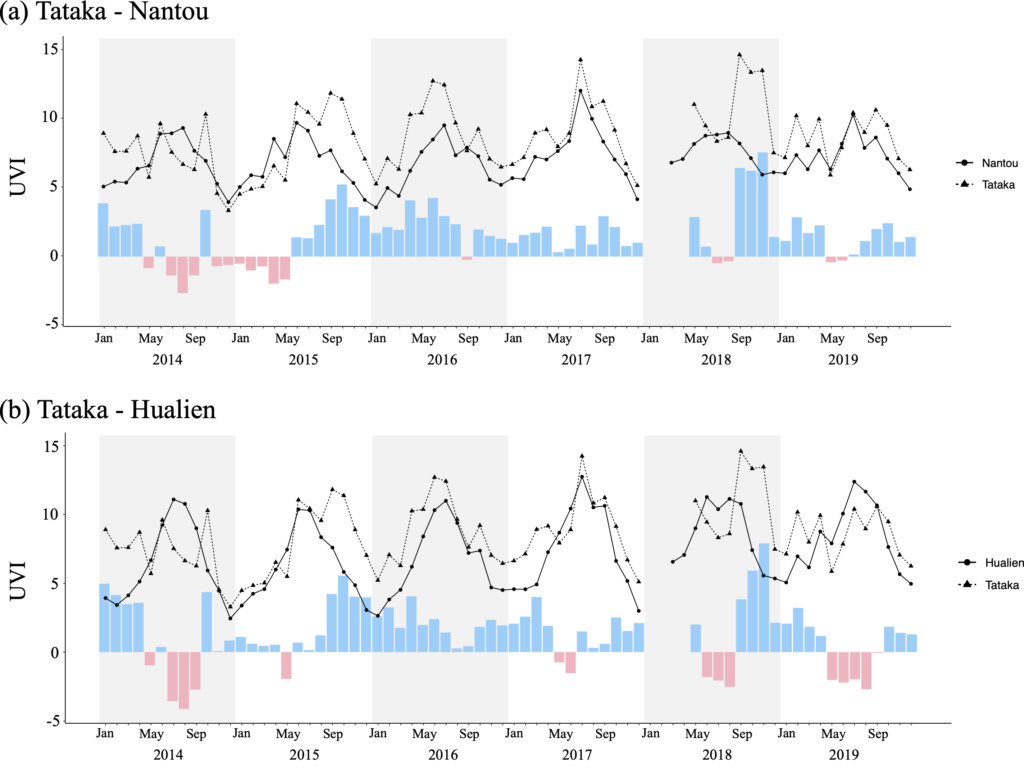
Figure 3 Monthly ultraviolet index (UVI) values for lowland and mountainous habitats of Rufous-capped Babblers from 2014 to 2019. The dashed line connects data from Tataka (a mountainous site) and the solid line connects data from (a) Nantou or (b) Hualien (two lowland sites). The bar plots show the differences of UVI (a) between Tataka and Nantou, and (b) between Tataka and Hualien with blue bars for positive values and red bars for negative values.
Our findings are consistent with the hypothesis that elevational differences in UV light environments can drive the evolution of plumage colouration. Different sexes and body parts respond to the plumage-associated selection forces differently. This study sheds new light on plumage evolution from a novel perspective—elevational divergence. Our findings also illustrate how ecological changes may drive phenotypic adaptation in organisms across a short distance. What amazes me most is that this evolutionary event has just happened along a mountain road, on which we often drive for a summer trip on this rugged island.
Image credit
Top right: A Rufous-capped Babbler caught in a high-altitude mountain habitat © Chih-Ming Hung.
References
Bertrand, J.A.M., Delahaie, B., Bourgeois, Y.X.C., Duval, T., García‐Jiménez, R., Cornuault, J., Pujol, B., Thébaud, C. & Milá, B. 2016. The role of selection and historical factors in driving population differentiation along an elevational gradient in an island bird. Journal of Evolutionary Biology 29: 824-836. VIEW
Dunn, P.O., Armenta, J.K. & Whittingham, L.A. 2015. Natural and sexual selection act on different axes of variation in avian plumage color. Science Advances 1: e1400155. VIEW
Gomez, D. & Théry, M. 2004. Influence of ambient light on the evolution of colour signals: Comparative analysis of a Neotropical rainforest bird community. Ecology Letters 7: 279-284. VIEW
Hill, G.E. & McGraw, K.J. 2006. Bird coloration: mechanisms and measurements. Vol. 1. Harvard University Press.
Johnson, A.E., Jordan Price, J. & Pruett-Jones, S. 2013. Different modes of evolution in males and females generate dichromatism in fairy-wrens (Maluridae). Ecology and Evolution 3: 3030-3046. VIEW
Medina, I., Langmore, N.E., Hall, M.L., Mulder, R.A., Cain, K.E., Peters, A. & Delhey, K. 2017. Habitat structure is linked to the evolution of plumage colour in female, but not male, fairy-wrens. BMC Evolutionary Biology 17: 1-9. VIEW
Stoddard, M.C. & Prum, R.O. 2008. Evolution of avian plumage color in a tetrahedral color space: a phylogenetic analysis of new world buntings. The American Naturalist 171: 755-776. VIEW

