

How do Broadbills produce that loud mechanical sound while displaying?
LINKED PAPER
Smithornis broadbills produce loud wing song by aeroelastic flutter of medial primary wing feathers. Clark, C.J., Kirschel, A.N.G., Hadjioannou, L. & Prum, R.O. 2016. Journal of Experimental Biology . DOI: 10.1242/jeb.131664 View
Non-vocal acoustic signals can be found in a number of avian lineages, and include percussive sounds such as those produced by club-winged manakins (Bostwick and Prum 2003), and flappet larks, aeroelastic flutter producing sounds such as those in hummingbirds (Clark et al. 2011) as well as sounds produced by other mechanisms including the whooshing sounds produced by flying pigeons and doves, and hornbills, among others (Clark and Prum 2015).
Another group of birds thought to produce sounds non-vocally as part of a visual display are Smithornis broadbills. Broadbills are Suboscine passerines, a group known for elaborate courtship displays, particularly those in the Manakin and Cotinga families, and elaborate and colourful plumage, again involving species in Cotingas and Manakins, but also species in the Old World suboscine families, Broadbills, Pittas and Asities. Compared to the colourful broadbills of Asia, Smithornis broadbills from Africa are relatively subdued in terms of plumage coloration (Fig. 1), but similarly to Manakins, perform an elaborate visual display, which has been presumed to function in mate attraction. During this visual display, they produce a very loud trill, which is thought to be mechanically-produced (Chapin 1953). However, the exact mechanism producing the sound had never been investigated, only that it was assumed that it was likely caused by modifications to the outer primary feathers (Chapin 1953).
Footage of Rufous-sided Broadbill (Smithornis rufolateralis) diplaying in Semliki National Park, Uganda © Alex Kirschel
During fieldwork in the Congo Basin in Africa in 2005 I was fortunate to witness a rufous-sided broadbill performing the display, and was fascinated by it. Subsequent visits to Uganda and Cameroon allowed me to watch the other species in the genus, African and Grey-headed broadbills, perform the display.

Figure 1 African broadbill (Smithornis capensis) photographed in October 2012, at Ngorongoro Crater, Tanzania. It has raised its white mantle feather, which are typically concealed, but are often prominent during the aerial display. At a distance in the dark understory, you might see a white flash moving in a tight circle © Alex Kirschel
I had put much thought into studying the sound-producing mechanism, but I lacked access to some of the necessary equipment, a high-speed camera and a wind-tunnel in particular. Then at ISBE in Perth in 2010 I discussed the study with Richard Prum from Yale, who shared my interest in the study and had the necessary equipment available. We agreed to collaborate and with his postdoc Chris Clark and a graduate student from the University of Cyprus, Louis Hadjioannou, we headed for the field. We went to Uganda in June 2011 to find two species of broadbills displaying in the jungle. After a few fruitless days we finally found our first African broadbills (S. capensis) displaying at Kibale National Park, and then rufous-sided broadbills (S. rufolateralis) at Semliki National Park. We obtained conventional 50 fps video using a Canon DSLR and high speed video using a Miro EX4 while recording the sound produced during the display using a Sound Devices digital recorder and Sennheiser microphone with a Telinga parabola. By comparing the high-speed videos with the recordings we identified a 1:1 correspondence between sound pulses and wingbeats in the display. We could also determine the onset of the sound during the downstroke of the wingbeat.
Chris Clark then ran experiments in a wind tunnel with individual primary feathers and a spread wing from specimens, and found that while any feather could be made to produce sound, P6 and P7 in particular, would flutter and produced sound at frequencies and airspeeds relevant to the broadbill’s aerial display.
Conventional and high-speed video footage of the display © Chris Clark, University of California Riverside
But we wanted to test this hypothesis. If P6 or P7 played such an important role in producing sound by fluttering during the display, what would happen to the sound if these feathers were manipulated? Louis and I returned to the field, this time to Tanzania, and performed some feather manipulation experiments on three individuals of African broadbill at three different sites. In each case manipulating the feather led to a change in the timbre of the sound, so that it was much less tonal, and more ratchety. Even when clipping 2 cm of the feather tip of either P6 or P7, we found tonality was reduced, and we established this quantitatively by comparing the entropy of the sound. Each manipulation led to a significantly higher entropy, meaning a less tonal sound was produced following feather manipulation (Fig. 2).
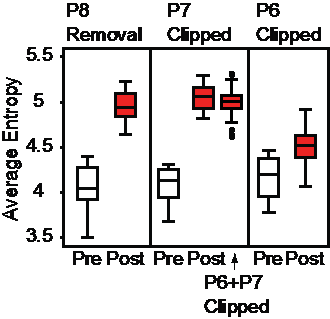 Figure 2 Manipulating feathers led to an increase in Average Entropy of the sound produced during the aerial display of African broadbill. This was found consistently, whether we removed P8, clipped 2 cm from the tip of P6, P7 or both. The role of P8 is less well understood, but it is thought that neighboring feathers may also play a role in how well sound-producing feathers can flutter to produce tonal sound. Adapted from Clark et al. (2016).
Figure 2 Manipulating feathers led to an increase in Average Entropy of the sound produced during the aerial display of African broadbill. This was found consistently, whether we removed P8, clipped 2 cm from the tip of P6, P7 or both. The role of P8 is less well understood, but it is thought that neighboring feathers may also play a role in how well sound-producing feathers can flutter to produce tonal sound. Adapted from Clark et al. (2016).
So why has this non-vocal sound producing mechanism evolved in Smithornis broadbills? This is a question that needs further investigation, but one hypothesis relates to their simplified syringeal morphology (Prum 1993). Smithornis lack the pessulus–a bony supporting element that separates the medial bronchial membrane into separate vibrating structures– and any intrinsic syringeal musculature (Prum 1993), but we don’t yet know if this simplified syringeal morphology precludes broadbills from producing loud vocal song. An alternative explanation is that non-vocal song may have evolved as a by-product of the visual flight display and led to subsequent simplification of syringeal morphology. The flight display and accompanying sound may have been preferred to vocal song by discerning females looking for mates.
References and further reading
Bostwick, K. S. & Prum, R. O. 2003. High-speed video analysis of wing-snapping in two manakin clades (Pipridae: Aves). J. Exp. Biol. 206: 3693-3706. View
Chapin, J. P. 1953. The birds of the Belgian Congo. Part 3. Bull. Am. Coll. Museum Nat. His. 75: 4-820. View
Clark, C. J. & Prum, R. O. 2015. Aeroelastic flutter of feathers, flight and the evolution of non-vocal communication in birds. J. Exp. Biol. 218: 3520-3527. View
Clark, C. J., Kirschel, A. N. G., Hadjioannou, L. & Prum, R. O. 2016. Smithornis broadbills produce loud wing song by aeroelastic flutter of medial primary wing feathers. Journal of Experimental Biology . 219: 1069-1075. View
Clark, C. J., Elias, D. O. & Prum, R. O. 2011. Aeroelastic flutter produces hummingbird feather songs. Science 333: 1430-1433. View
Gorman, J. 2016. For African Broadbills, Seduction to the Sound of Feathers. The New York Times. View
Pennisi, E. 2016. Video: African bird ‘sings’ by fluttering its wings. Science. View
Prum R.O. 1993. Phylogeny, biogeography, and evolution of the broadbills (Eurylaimidae) and asities (Philepittidae) based on morphology. Auk 110: 304-324. View
Image credit
Featured image: African Broadbill, Smithornis capensis © Francesco Veronesi
If you want to write about your research in #theBOUblog, then please see here.

