

LINKED PAPER
The reliability of achromatic displays is island-dependent in nocturnal Storm Petrels. Soravia, C., Aguado-Giménez, F. & Avilés, J.M. 2019. IBIS. DOI: 10.1111/ibi.12772. VIEW
“Up above the sea’s grey flatland, wind is gathering the clouds. In between the sea and clouds proudly soaring the Petrel, reminiscent of black lightning” (The song of the Stormy Petrel, Maxim Gorky, 1901). For the Russian writer Maxim Gorky, the the European Storm Petrel Hydrobates pelagicus was the free spirit that welcomes the storm of revolution. Before him, it was the bearer of bad weather for sailors. Its habit of foraging in open sea carried by the wind and its frequent nocturnal activity, especially during breeding, gives the Storm Petrel its evocative nature. However, upon closer inspection, this “proud black demon of the tempest” appears far less ominous, with less than 30 g of body mass, and it is surprisingly not black.
Storm Petrels are covered in dark plumage, with the exception of white bands on the underwings and the rump. We examined chest and rump feathers of over 180 Storm Petrels with UV-visible reflectance spectrometry, discovering that the feathers composing the white band on the rump are highly reflective in the ultraviolet (UV, 300-400 nm) (Fig 1a). Interestingly, the reflectance spectrum of black chest feathers does not follow the pattern typical of melanins, but shows a reflectance peak both in the UV and in the yellow-red wavelengths (550-700 nm) (Fig 1b), which accounts for its shiny appearance and brown shades. The colour of rump and chest feathers also does not differ in males and females.
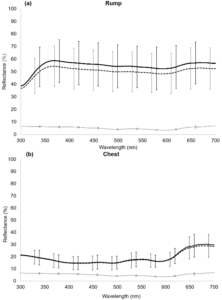 Figure 1 Mean reflectance spectrum (mean ± SD) of (a) rump feathers and (b) chest feathers (males: solid line, females: dashed line). In (a) and (b), the grey line represents the reflectance spectrum of the black background paper used for measurements
Figure 1 Mean reflectance spectrum (mean ± SD) of (a) rump feathers and (b) chest feathers (males: solid line, females: dashed line). In (a) and (b), the grey line represents the reflectance spectrum of the black background paper used for measurements
The white, UV reflective rump band must appear very conspicuous against the darker surrounding plumage and the environmental background, especially during nocturnal courtship displays, when mates chase each other over the nesting burrow. As underlined by zoologist Peter Davis in the 1950s, the rump band seems “designed to elicit a visual response of some kind” (Davis 1957). The use of visual communication in species with nocturnal habits has been considered negligible until recently because chromatic discrimination is impaired at low lights (Kelber et al. 2003). However, white and dark plumage patterns can constitute effective achromatic displays (Penteriani & Delgado 2017). So, over sixty years after Davis’ observation, it is time to address the question: can Storm Petrels’ plumage coloration act as a signal?
The answer is yes – a browner chest and more intense rump UV coloration were associated with higher body mass (corrected for tarsus length) in breeding males, but not in females. Male plumage coloration can thus signal individual condition to the partner during courtship displays or to rivals when defending the nest. Here comes the interesting part though: the strength of the colour-condition relationship varies in different breeding islands, in fact, in one island the relationship was absent.
The study population breeds on three small islands along the coast of Murcia region (Southern Spain) (Fig 2): Isla de las Palomas, Isla Grosa, and Isla Hormiga. The first two islands, literally Island of the Doves and Awesome Island, sound more inviting than the third, Ant Island. Quite the contrary, the first two islands, Grosa and Palomas, are close to the coast (0.8-2.5 km), exposed to tourism, predation, and interspecific competition for nest sites. Instead, Isla Hormiga is distant from the coast (4 km), part of an integral marine reserve, and the only potential predators on the island are about 20 pairs of Yellow-legged Gulls Larus michahellis, so it offers a more favourable breeding environment. The different exposure to predation and human disturbance between islands actually affects individuals’ condition: males from Isla Hormiga are heavier than males from Isla Grosa and Isla de las Palomas. The likely reason is that human disturbance is perceived as predation risk (Watson et al. 2014) and adds to direct predation risk, altering hormonal profiles (Zanette et al. 2014, Soldatini et al. 2015) and foraging behaviour (Mougeot & Bretagnolle 2000, Rubolini et al. 2015).
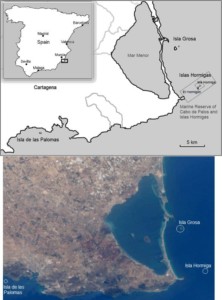 Figure 2 Map of the location of the three islands: Isla Grosa, Isla de las Palomas, Isla Hormiga. Satellite image modified from: Image Science & Analysis Laboratory, Johnson Space Center, NASA| Wikimedia Commons
Figure 2 Map of the location of the three islands: Isla Grosa, Isla de las Palomas, Isla Hormiga. Satellite image modified from: Image Science & Analysis Laboratory, Johnson Space Center, NASA| Wikimedia Commons
If plumage coloration depends on male condition, we would predict that only the best males will produce intense chest and rump coloration. However, this is not necessarily true. If resources are abundant and easily available, even males of poorer quality could afford to invest in a colour signal (Vergara et al. 2012). We believe that this is the case in our study population: neither chest nor rump coloration were related to male body mass in Isla Hormiga (Fig 3), where the favourable conditions may relieve males from the need to trade-off the investment between their body maintenance and the production of colour signals. On the contrary, both chest-yellow red chroma, i.e. colour saturation in the yellow-red wavelengths, and rump UV chroma positively predicted male body mass in Isla Grosa and Isla de las Palomas (Fig 3).
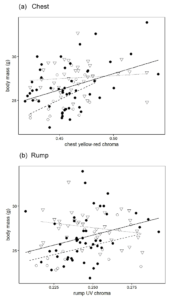
Figure 3 Relationship between male body mass and (a) chest yellow-red chroma (b) rump UV chroma according to the island of origin (Isla de las Palomas: filled dots, solid line; Isla Grosa: empty dots, dashed line; Isla Hormiga: triangles, dotted line)
Therefore, the achromatic plumage pattern of Storm Petrels, “reminiscent of black lightning“, does have the potential to act as a signal of the condition of the bearer, but the information conveyed is not always true. This study shows that the reliability of a signal can vary with environmental conditions and we should not give it for granted when studying visual communication and sexual selection in wild animal populations.
Nominate this article for a BOU Science Communication Award.
References
Beauchamp, G. & Heeb, P. 2001. Social foraging and the evolution of white plumage. Evol. Ecol. Res. 3: 703-720. VIEW
Davis, P. 1957. The breeding of the Storm Petrel. British Birds 50: 85-101. VIEW
Gorky, M. 1901. The Song of the Stormy Petrel. In: Gorky, Maksim; Sukennikov, M. 1902. Tri razskaza (Three Stories), Izd-vo Ioanna Rėde, pp. 20–26. VIEW
Kelber, A., Vorobyev, M. & Osorio, D. 2003. Animal colour vision – behavioural tests and physiological concepts. Biol. Rev. 78: 81-118. VIEW
Mougeot, F. & Bretagnolle, V. 2000. Predation risk and moonlight avoidance in nocturnal seabirds. J. Avian Biol. 31: 376-386. VIEW
Rubolini, D., Maggini, I., Ambrosini, R., Imperio, S., Paiva, V.H., Gaibani, G., Saino, N. & Cecere, J.G. 2015. The effect of moonlight on Scopoli’s Shearwater Calonectris diomedea colony attendance patterns and nocturnal foraging: a test of the foraging efficiency hypothesis. Ethology 121: 284-299. VIEW
Penteriani, V. & Delgado, M.M. 2017. Living in the dark does not mean a blind life: bird and mammal visual communication in dim light. Phil. Trans. R. Soc. B 372: 20160064. VIEW
Vergara, P., Martínez‐Padilla, J., Mougeot, F., Leckie, F. and Redpath, S.M. 2012. Environmental heterogeneity influences the reliability of secondary sexual traits as condition indicators. J. Evol. Biol. 25: 20-28. VIEW
Watson, H., Bolton, M. & Monaghan, P. 2014. Out of sight but not out of harm’s way: human disturbance reduces reproductive success of a cavity-nesting seabird. Biol. Cons. 174: 127-133. VIEW
Image credit
Featured image: Mediterranean Storm Petrel Hydrobates pelagicus melitensis © Felipe Aguado-Giménez

