


LINKED PAPER
Artificial light at night affects the timing of roosting by Chimney Swifts. Dougherty, E., Parzer, H. F., Morton, E. R. 2024. IBIS. DOI: 10.1111/ibi.13296. VIEW
The negative impacts of humans on wildlife are widely accepted to be driving many species towards extinction. While there are ongoing efforts to reduce or reverse these impacts, in order to do this effectively we must first understand how animals are being affected. Light pollution is an increasingly widespread phenomenon shown to affect numerous organisms which rely on natural light to maintain and adjust their biological rhythms, but there are still lots of questions to be answered about how changing light availability can impact the physiology and behaviour of different species.
In a recent study in Ibis, Emma Dougherty and colleagues examined the effect of artificial light at night (ALAN) on roosting behaviour of Chimney Swifts (Chaetura pelagica), to investigate whether light pollution may be a relevant factor influencing this species.
Communal roosting migratory birds
Chimney Swifts are small, diurnal, migratory, aerial insectivores listed as Vulnerable by the IUCN (Lowen et al. 2019). The species roost communally in large vertical cavities, almost exclusively using artificial structures such as chimneys (Zammuto & Franks 1981, Zanchetta et al. 2014). The size of the roosting group changes seasonally, but year-round the initiation of roosting is a synchronised event and all swifts enter within a short period of time around sunset, with variation depending on season, temperature, and wind conditions (Michael & Chao 1973).
In this study, 21 roosting sites were identified in Northern New Jersey and the New York Metropolitan area and observed between June 2021 and October 2022 during the swifts’ period of breeding and post-breeding migration. The researchers used generalised linear mixed models (GLMMs) to test the hypothesis that light pollution affected the timing of roosting. The response variables included the start and end time of entry relevant to sunset, while fixed variables were estimated light pollution, number of birds, temperature, month, relative humidity, and wind speed. Roosting site was included as a random effect to account for a lack of independence across multiple observations.
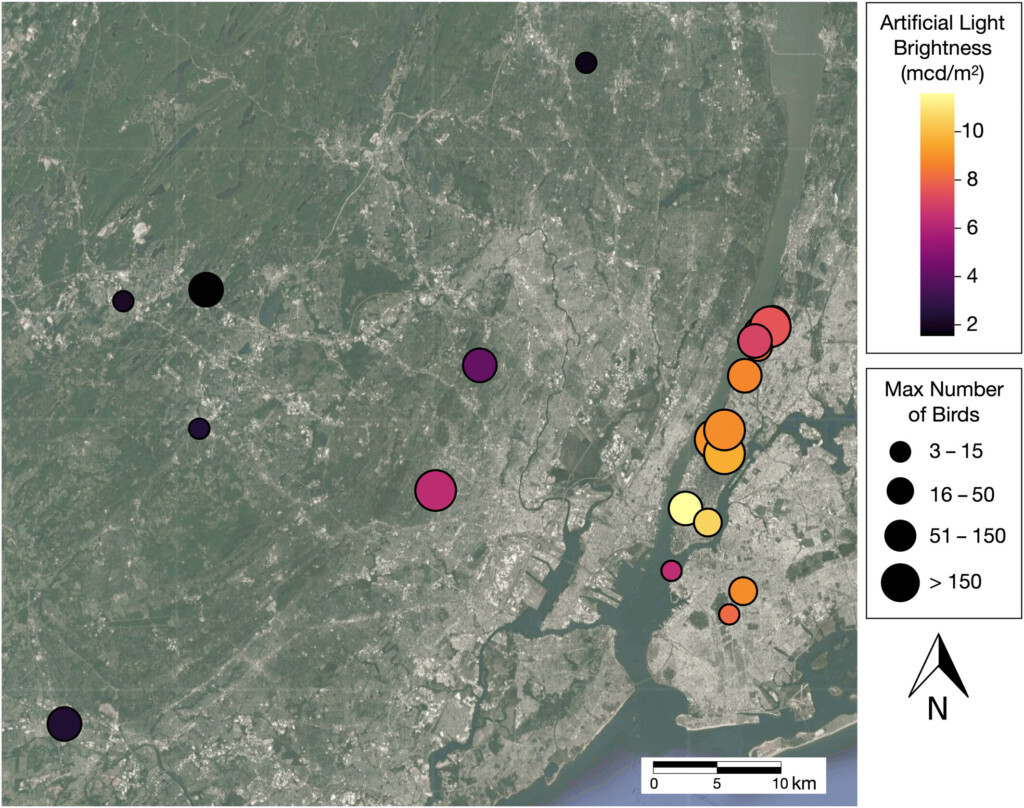
Figure 1. Map of Chimney Swift roosting sites in the North New Jersey and New York Metropolitan Area. Points indicating site locations are colour-coded based on artificial light brightness (low light pollution = black; high light pollution = yellow (colour) or white (greyscale)) and point size corresponds to the maximum number of birds observed at each site.
Light pollution significantly affects Chimney Swifts
The results showed that ALAN significantly affected the entry behaviour of communal roosting Chimney Swifts, with swifts entering roosts exposed to elevated levels of light pollution significantly later relative to sunset compared with roosts with less exposure. Also, in areas of high light pollution the swifts did not initiate roosting earlier in the summer months compared with autumn, as they did in areas with low light pollution.
The delayed roost entry with increased ALAN may be due to the artificial light overriding solar stimuli used by swifts as a cue to enter roosts (Michael & Chao 1973). However, it is also possible that swifts stay out longer if ALAN provides additional opportunities to catch insects successfully, increases predator detection through improved visibility, or induces a direct or indirect stress response. Further studies could elucidate whether any of these factors act as drivers of the delay.
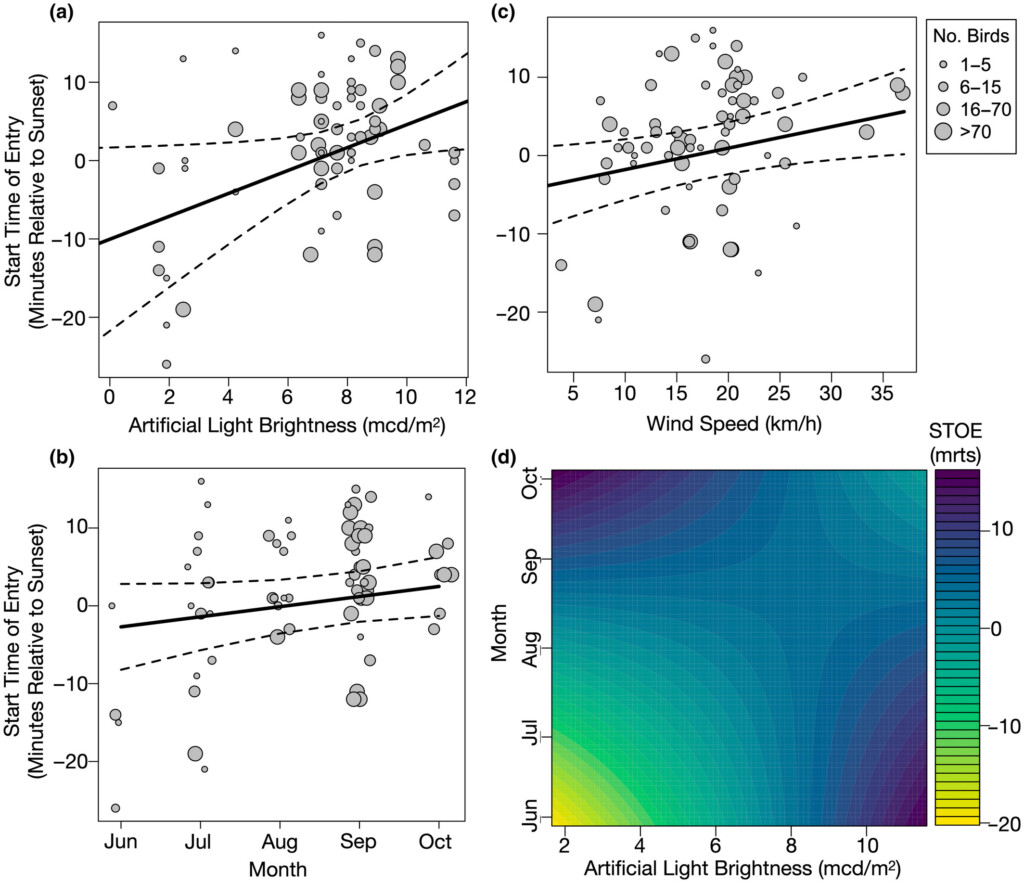
Figure 2. Predicted relationships between start time of entry into roosting sites and (a) artificial light brightness (light pollution), (b) month, (c) wind speed, and (d) the interaction between light pollution and month based on the most parsimonious generalized linear mixed model. In (a)–(c) each point represents an observation and larger points indicate more Chimney Swifts. The lines of best fit (solid lines) and 95% confidence intervals (dashed lines) are indicated. In (d) colour ranges from yellow (colour) or white (greyscale), representing an earlier start time of entry (STOE) to black, representing a later start time of entry. Entry times are minutes relative to sunset (mrts).
Regardless of the cause, delayed roost entry with increased ALAN likely makes swifts more vulnerable to predators. In the Northern hemisphere, a variety of birds of prey have been shown to utilise ALAN to catch their prey (e.g. Kettel et al. 2016). Another potential consequence of ALAN could be an impact on the swifts’ migratory behaviour, as has been found in other species (Smith et al. 2021).
The findings also showed that Chimney Swifts began roosting later prior to post-breeding migration compared with during the breeding season, potentially driven by changes in insect availabilities, and that both wind and group size had an effect on timing of roost entry. Birds were shown to enter the roosts later in windy conditions, potentially due to increased physical difficulty. Larger groups were found to enter roosts earlier than smaller groups, which may be due to larger groups needing to start entering earlier because it takes them longer overall, however this effect was only found in areas of low ALAN.
Despite the success of Chimney Swifts in taking advantage of anthropogenic structures following the destruction of their native habitat (large hollow trees) through widespread deforestation, the results of this study clearly indicate that they are now being impacted by human-induced light pollution. As this already threatened species is continuing to decline, understanding the potential consequences of ALAN could go towards helping us better assess the threats it is facing, and ultimately help to determine the best conservation strategies to put in place.
References
Kettel, E.F., Gentle, L.K. & Yarnell, R.W. (2016). Evidence of an urban Peregrine Falcon (Falco peregrinus) feeding young at night. Journal of Raptor Research 50: 321–323. VIEW
Lowen, J., Wheatley, H., Hermes, C., Burfield, I. & Wege, D. (2019). The 2018 IUCN red list in the neotropics. Neotropical Bird 24: 48–54. VIEW
Michael, E.D. & Chao, W.T.H. (1973). Migration and roosting of Chimney Swifts in East Texas. The Auk 90: 100–105. VIEW
Smith, R.A., Gagné, M. & Fraser, K.C. (2021). Pre-migration artificial light at night advances the spring migration timing of a trans-hemispheric migratory songbird. Environmental Pollution 269: 116136. VIEW
Zammuto, R.M. & Franks, E.C. (1981). Environmental effects on roosting behavior of Chimney Swifts. The Wilson Bulletin 93: 77–84. VIEW
Zanchetta, C., Tozer, D., Fitzgerald, T., Richardson, K. & Badzinski, D. (2014). Tree cavity use by Chimney Swifts: Implications for forestry and population recovery. Avian Conservation and Ecology 9: 1. VIEW
Image credits
Top right: Chimney Swift (Chaetura pelagica) | Andrew C. | CC BY 2.0 Wikimedia Commons
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here

