


Forest passerine survival rates go down with increasing latitude
LINKED PAPER
Age effects on survival of Amazon forest birds and the latitudinal gradient in bird survival. Muñoz, A. P., Kéry, M., Martins, P. V. & Ferraz, G. 2018. The Auk. DOI: 10.1642/AUK-17-91.1. VIEW
Bold, unexplained patterns are fascinating. The consistency of species-area relationships, population cycles, and allometric rules, has motivated biologists for many years and pages of literature. For biologists living in the tropics, perhaps the most striking of these patterns are the various latitudinal gradients in community and life history metrics. Science excites our curiosity in the search for universal rules, but large-scale, latitudinal changes create the appearance that some things may be fundamentally different in different parts of the world. Could there be really such a thing as ‘tropical biology’?
Perhaps the best-documented latitudinal gradient in life history is the tendency for avian clutch size to increase with increasing latitude, both at the intra-specific and interspecific levels, in the old and the new world, in the northern and southern hemispheres. Since there is no evidence that high-latitude populations grow faster than equatorial ones, it has been widely understood that some other life-history trait must also change with latitude, to compensate for the variation in clutch size. Thus, throughout the second half of the 20th century, the idea that adult tropical birds live longer than their temperate counterparts became widely accepted among ornithologists.
 Figure 1 A Guianan Warbling Antbird, Hypocnemis cantator, banded and ready for release © Erik Johnson
Figure 1 A Guianan Warbling Antbird, Hypocnemis cantator, banded and ready for release © Erik Johnson
The acceptance of a latitudinal gradient in survival, however, was more based on common sense than hard data. So much so, that when Karr et al. (1990) challenged the idea in a paper subtitled “Will the dogma survive?” there were no voices sounding in defense of the “dogma”. Indeed, two decades later, Corey Tarwater and colleagues suggested that, if there is no latitudinal trend in the survival of adult birds, there could be a trend in the survival of juveniles (Tarwater et al. 2011). Their work drew on detailed analysis of age-dependent survival in one Central American passerine species and suggested that tropical juveniles would survive more than temperate juveniles while adult survival would show no relationship with latitude. This would imply, of course, relatively little difference between adult and juvenile survival in the tropics.
In a paper recently published in The Auk, we use bird-banding data to tackle the problem of the latitudinal gradient in bird survival on two fronts: first, we quantify the effect of age on survival for forty species of Amazon forest passerines and find that adults of their tropical site have substantially higher survival probabilities than juveniles (Fig. 2); second, we analyse 342 survival estimates from 175 species in twelve regions from Peru to Alaska and find that survival does go down with increasing latitude (Fig. 3). The latitudinal effect persists after accounting for effects of migration mode, phylogeny, and time of data collection. Roughly speaking, the odds of annual survival at the Equator are nearly 1.7 times as high as at 60º latitude. We conclude that the latitudinal gradient in survival is a fact, at least as seen among New World forest passerines. The “dogma” need no longer be seen as a dogma.
From the data-collection perspective, our work benefited a great deal from two recent advances. First, there is an ongoing transformation in how tropical ornithologists assess bird age. The Wolfe-Ryder-Pyle molt-cycle system (Wolfe et al. 2010, 2012), which we employed, is extremely useful for aging birds in populations that have poorly delimited breeding periods, which is typical of tropical regions. This was central for quantifying the effect of age on survival. Second, we tapped into the phenomenal database of Vital Rates of North American Birds made available by the Institute of Bird Populations (IBP). This source enabled them to analyse estimates from as far North as Alaska. The IBP estimates, combined with a wealth of tropical passerine survival estimates published since 1990, made it possible to take up Karr et al.’s (1990) challenge anew, with a substantial amount of data.
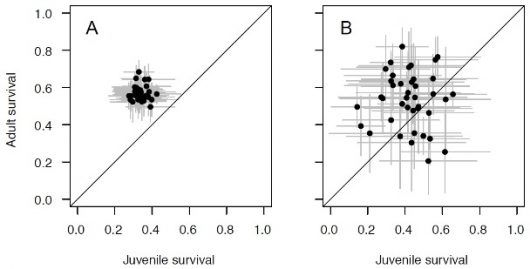 Figure 2 Adult and Juvenile survival probability estimates for forty Amazon forest passerine species. The two panels show results from two different statistical approaches that offer qualitatively similar results. The approach on the left (A) treats species as being individual samples of a wider set of species whose distribution of survival probability we want to characterize; the approach on the right (B) analyzes every species on its own, independently of all the others. Gray lines represent standard deviations of the estimated survival. In both cases it adults tend to survive more than juveniles
Figure 2 Adult and Juvenile survival probability estimates for forty Amazon forest passerine species. The two panels show results from two different statistical approaches that offer qualitatively similar results. The approach on the left (A) treats species as being individual samples of a wider set of species whose distribution of survival probability we want to characterize; the approach on the right (B) analyzes every species on its own, independently of all the others. Gray lines represent standard deviations of the estimated survival. In both cases it adults tend to survive more than juveniles
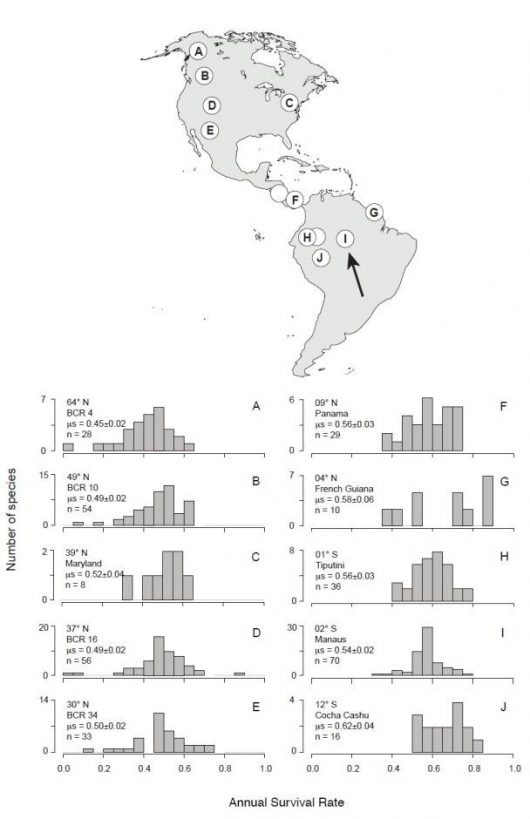 Figure 3 Latitudinal change in the distribution of survival rates. The map shows the twelve regions from where we compiled survival estimates, including our own study area near Manaus, Brazil, at the tip of the black arrow. The two blank circles in Ecuador and Costa Rica indicate locations with only one survival estimate each, not shown on histograms at the right-hand side of the figure. Histograms show the number of species per annual survival rate class, with text indicating the latitude, region name, mean and standard deviation of the region’s survival rate and the number of species sampled in each region. ‘BCR’ stands for ‘Bird Conservation Region’ in the Institute of Bird Populations’ Vital Rates of North American Landbirds database
Figure 3 Latitudinal change in the distribution of survival rates. The map shows the twelve regions from where we compiled survival estimates, including our own study area near Manaus, Brazil, at the tip of the black arrow. The two blank circles in Ecuador and Costa Rica indicate locations with only one survival estimate each, not shown on histograms at the right-hand side of the figure. Histograms show the number of species per annual survival rate class, with text indicating the latitude, region name, mean and standard deviation of the region’s survival rate and the number of species sampled in each region. ‘BCR’ stands for ‘Bird Conservation Region’ in the Institute of Bird Populations’ Vital Rates of North American Landbirds database
From the analytical perspective, we used a multi-species Cormack-Jolly-Seber (CJS) approach to model our Amazon data. The CJS is a standard method for estimating apparent survival from capture-recapture data on open populations. The ‘multi-species’ approach means we treat each of the forty species in our data as a random draw from a wider distribution of species that includes our sample. The inference about age effects was made at the level of this wider distribution, strengthening the generality of the conclusions. The CJS accounts for the possibility of capture failure, and a ‘mixture’ component of the model, developed by co-author Marc Kéry, makes it possible to incorporate birds of unknown age in the analysis. The work thus addresses a variety of sources of uncertainty before reaching its final conclusions.
 Figure 4 An adult White-plumed Antbird, Pithys albifrons © Pedro Martins
Figure 4 An adult White-plumed Antbird, Pithys albifrons © Pedro Martins
The observation of a latitudinal trend in survival probability helps understand the maintenance of a latitudinal trend in clutch size, but it need not be the only explanation. It is also possible, for example, that the number of clutches laid each year also vary with latitude in association with the length of the breeding season. With or without latitudinal change in number of clutches, one cannot tell whether the variation in survival documented in our paper is a sufficient explanation for the current variation in clutch size. Furthermore, even if one explains the maintenance of current variation, there is still the question of how that variation evolved.
There is certainly much to explore in the latitudinal variation of bird life history traits. It is striking that a relatively high survival probability is found in regions where the bird ‘pace of life’ is relatively slow, as measured by their basal metabolic rate (Wiersma et al. 2007). Intuitively, ‘slow pace’ is coherent with ‘long-lived’, but why would tropical birds, which inhabit relatively hot environments, have a particularly slow basal metabolic rate? Unless there isn’t a straightforward positive relationship between metabolic rate and temperature (Clark & Fraser 2004). Challenging evolutionary and physiological questions remain to be answered and hopefully will motivate further study of this puzzling latitudinal trend.
References
Clarke, A. & Fraser, K.P.P. 2004. Why does metabolism scale with temperature. Funct. Ecol. 18: 243-251. VIEW
DeSante, D.F., Kaschube, D.R. & Saracco, J.F. 2015. Vital rates of North American landbirds. VIEW
Karr, J.R., Nichols, J.D., Klimkiewicz, M.K. & Brawn, J.D. 1990. Survival rates of birds of tropical and temperate forests: will the dogma survive? Am. Nat. 136: 277-291. VIEW
Muñoz, A.P., Kéry, M., Martins, P.V. & Ferraz, G. 2018. Age effects on survival of Amazon forest birds and the latitudinal gradient in bird survival. Auk 135: 299-313. VIEW
Tarwater, C.E., Ricklefs, R.E., Maddox, J.D. & Brawn, J.D. 2011. Pre-reproductive survival in a tropical bird and its implications for avian life histories. Ecology 92: 1271-1281. VIEW
Wiersma, P., Muñoz-Garcia, A., Walker, A. & Williams, J.B. 2007. Tropical birds have a slow pace of life. PNAS 104: 9340-9345. VIEW
Wolfe, J.D., Ryder, T.B. & Pyle, P. 2010. Using molt cycles to categorize the age of tropical birds: an integrative new system. J. Field Ornithol. 81: 186-194. VIEW
Wolfe, J.D., Ryder, T.B., Pyle, P. & Johnson, E.I. 2012. Using molt and plumage cycles to age tropical birds: updates and recent advances. Ornitol. Neotrop. 23: 153-158. VIEW
Image credit
Featured image: A Rufous-throated Antbird, Gymnopithys rufigula © Erik Johnson
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here.

