

LINKED PAPER
Context‐dependent conservation of the cavity‐nesting European Roller. Finch, T., Branston, C., Clewlow, H., Dunning, J., France, A.M.A., Račinskis, E., Schwartz, T. & Butler, S. 2019. IBIS. DOI: 10.1111/ibi.12650. VIEW
Conservationists are often tasked with identifying which factors limit the populations of threatened species, so that action can be taken to turn things around. Text-book examples of this sort of evidence-informed conservation success include the provisioning of food-rich habitat by farmers in south-west England to aid the recovery of Cirl Buntings, Emberiza cirlus, and the installation of artificial nesting sites in Portugal to support Lesser Kestrels, Falco naumanni.
Generally, breeding populations are limited by the availability of one of two key functional resources: foraging habitat or nesting habitat. Yet, nesting habitat can only be limiting whilst foraging habitat is sufficient, and vice versa. If Lesser Kestrel numbers were to continue to grow, following the installation of more and more artificial nest-sites, then eventually they’d hit a ceiling determined by foraging resource availability. This raises the interesting possibility that different factors might be limiting populations in different parts of a species’ range. If so, then conservation science conducted in one place might not be generalisable to others.
The European Roller, Coracias garrulus, has much in common with the Lesser Kestrel; they’re similar in size, eat mainly insects, and nest in cavities (but can’t excavate their own). Conservationists are concerned about the European Roller, and artificial nest-boxes have been used with some success; for example, schemes in parts of France (Aleman & Laurens 2013), Serbia (Ružić et al. 2014) and Hungary (Molnar 1998) have led to some encouraging population increases. But not all nest-box schemes have been successful; Rollers are just about hanging on in Poland (Sosnowski & Chmielewski 1996) and went extinct several years ago in Estonia (Lüütsepp et al. 2011) despite the presence of unoccupied and apparently suitable nest-boxes.
As part of my PhD – with lots of help – I monitored two Roller nest-box schemes in two very different parts of Europe. The French site is dominated by vineyards, orchards and fallows. There aren’t many trees, but some Rollers make use of old Iberian Green Woodpecker, Picus sharpei nests in lines of Poplar and Cypress hedges; most nest in the wooden nest-boxes adorning every other telegraph pole, with a handful nesting in old buildings. 2200 km to the north east, the Latvian site is on an ancient sand dune system, covered in sparse Pine forest and open heath. Here, a handful of Rollers use old Black Woodpecker, Dryocopus martius nests in Pine trees, but, as in France, most are now in nest-boxes.
A large part of my fieldwork involved counting insects – potential Roller prey (Fig. 1) – in different land cover types. This allowed me to produce a map of estimated foraging resource availability across each study site. Perhaps unsurprisingly, given the climatic differences, the average biomass of insects per unit area was 5 times higher in France (mostly grasshoppers and cicadas) than in Latvia (grasshoppers and some tasty-looking, but thinly-spread, beetles). Perhaps as a result of this, the density of nesting Rollers was 4 times higher in France, with 42% of nest-boxes occupied compared to just 13% in Latvia (the overall density of nest-boxes was similar between the two sites).

Figure 1 Some potential Roller prey: (a) a Bush Cricket (possibly Tettigonia viridissima), and (b) a Dor Beetle (family Geotrupidae). (c) shows a large, insect-rich fallow at the French site. (d) shows a nest-box fixed to a Pine tree in Latvia. The plastic sheath around the trunk is intended to stop Pine Martens climbing up © Tom Finch
More interesting, I think, was the spatial distribution of breeding pairs within each study area. In Latvia, we found a convincing relationship between nest-box use and foraging resource availability, with territories clustered around the open, sunnier habitats where insect activity is greatest (Fig. 2). We found no such pattern in France, where instead Rollers territory density was simply correlated with nest-box density (this was not the case in Latvia).
What this suggests is that, at the French Roller population is still below the ceiling set by foraging resource availability. If the entire study area was as well served with nest-boxes as the most densely-served 1-km square (ca. 16 boxes, compared to an average of 5.4 per km2), then the entire study area might perhaps be as densely populated by Rollers as this square (ca. 10 pairs, compared to an average of 2.2 per km2). In Latvia, however, it seems that there are more than enough nest-boxes, but that the population is limited by foraging resource availability; any population increase will likely require habitat management, rather than additional nest-boxes.
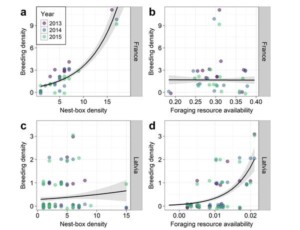
Figure 2 Local breeding density correlates with nest-box density in France (a) but not Latvia (c), and with estimated foraging resource availability in Latvia (d) but not France (b). Each point represents one 1‐km square. Dashed lines and shaded regions represent predictions (±se) of the best‐fitting model
This isn’t to say that nest-boxes aren’t important in Latvia; given the limited availability of old Black Woodpecker holes, competition with Stock Doves, Columba oenas, and predation by Pine Martens, nest-boxes have almost certainly saved this isolated population from extinction. Equally, any reduction in foraging resource availability in France might mean that nest-box occupation starts to fall. But our study does suggest that, at present, the priorities for Roller conservation might differ across its European breeding range.
Two caveats to end with. Approximately 12% of nest-boxes at our French site had damaged or missing lids, and these boxes were twice as likely to remain empty compared with intact boxes. Population gains associated with nest-boxes will therefore be short-lived unless broken boxes are repaired or replaced. Second, the consequences of using nest-boxes rather than natural cavities are poorly understood. Evidence from Portugal suggests that nest-boxes do a poor job of insulating nests against temperature extremes (Catry et al. 2015). It’s also possible that conspicuous nest-boxes expose Rollers to elevated levels of competition or predation (Rodríguez-Ruiz et al. 2011).
References
Aleman, Y. & Laurens, J. 2013. Répartition et effectifs du Rollier d’Europe (Coracias garrulus) dans les Pyrénées-Orientales en 2011. La Mélano 13: 1-11.
Catry, I., Catry, T., Patto, P., Franco, A.M.A., Moreira, F. 2015. Differential heat tolerance in nestlings suggests sympatric species may face different climate change risks. Climate Research 66: 13-24. VIEW
Lüütsepp, G., Kalamees, A. & Lüütsepp, O. 2011. European Roller Coracias garrulus in Estonia 2000–2011. Hirundo 24: 61-72.
Molnár, G. 1998. Breeding biology and foraging of Rollers (Coracias garrulus) nesting in nest-boxes. Ornis Hungarica 8 Suppl. 1: 119-124.
Rodríguez-Ruiz, J., Avilés, J.M. & Parejo, D. 2011. The value of nestboxes in the conservation of Eurasian Rollers Coracias garrulus in southern Spain: Nestboxes and Roller conservation. IBIS 153: 735-745. VIEW
Ružić, M.M., Szekeres, O., Ágoston, A., Balog, I. & Brdarić, B. 2014. The recovery of the European Roller Coracias garrulus population in Vojvodina Province, Serbia. Proceedings of the Second Adriatic Flyway Conference in Durrës, Albania.
Sosnowski, J. & Chmielewski, S. 1996. Breeding biology of the Roller Coracias garrulus in Puszcza Pilicka Forest (Central Poland). Acta Ornithologica 31: 119-131. VIEW
Image credits
Featured image: European Roller, Coracias garrulus © Tom Finch

