

LINKED PAPER
Male courtship preference during seasonal sympatry may maintain population divergence. Kimmitt, A.A., Dietz, S.L., Reichard, D.G. & Ketterson, E.D. 2018. Ecology and Evolution. DOI: 10.1002/ece3.4640. VIEW
Seasonal bird migration has long fascinated humans because of its taxing nature: in order to survive, birds must undergo physiologically demanding transitions to tackle long journeys to more suitable habitat. But one question scientists continue to ponder is: how is there so much diversity within migratory species? If birds move around a lot, wouldn’t they be able to easily breed across their ranges and reduce diversification? A recent study surprisingly found that migratory bird lineages have diversified more than their sedentary counterparts (Rolland et al. 2014). One focus of the Ketterson lab’s research is to understand one of the ways that variation in migratory strategies can promote divergence.
Within migratory species, populations often exhibit a spectrum of migratory behavior from sedentary (or resident) populations to obligate long-distance migrants. Within these species complexes, migrants and residents can exhibit interesting distributions by geographically overlapping at different times of year. For example, seasonal sympatry (also known as heteropatry) is when closely related migrant and resident populations are found in the same geographic space typically in the winter and early spring; following the spring migration, populations are then geographically isolated (Winker 2010). Residents may begin breeding prior to departure of the migrants. This presents an opportunity for gene flow between the populations; however, differences in breeding timing and/or population-specific mating preferences could prevent interbreeding and maintain speciation (Ketterson et al. 2015).
In this recent study, we investigated mating preferences in Dark-eyed Juncos, Junco hyemalis, a small songbird that exhibits seasonal sympatry. Populations differ in bill coloration and also have slight differences in plumage coloration, wing shape and size, in addition to differences in migratory behavior. Resident juncos live year-round in the Appalachian Mountains and are joined by migrant juncos from October through April. Residents often begin breeding in early April, and migrants and residents have exhibited differences in breeding timing. Reproductive residents, therefore, are encountering other juncos that exhibit varying levels of reproductive readiness.
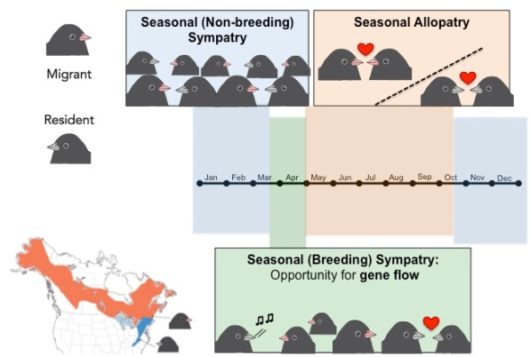
Figure 1 Resident and migrant juncos are found in seasonal sympatry during the winter months and early breeding season (see blue on map). Migrants and residents return to seasonal allopatry (i.e. populations are geographically isolated) after the spring migration when migrants return to their breeding grounds (see orange on map) © Figure adapted from Fudickar et al. 2016; designed by J.W. Atwell.
Past research has put an emphasis on female mate choice, because females typically invest more in reproduction, whereas males are limited by their number of mates. But dark-eyed juncos are socially monogamous, meaning that males and females form pair bonds and both invest in caring for the offspring for the duration of the breeding season. Males also typically enter reproductive condition prior to females (Williams 2012), as they have to establish and defend a territory before finding a mate. So as resident males begin courting females at the start of the breeding season, how do they avoid wasting time courting a non-reproductive mate? Especially one that will abandon during spring migration?
We presented free-living male juncos with either a migrant or resident caged female accompanied by a recording of a population-specific female vocalization. When the male came within 10 meters of the female, we recorded all of the male’s courtship behaviors. When a male junco courts a female, he puffs out his body feathers (i.e. ptiloerection), spreads his tail to show off flashy white tail feathers, and sings a specific courtship song, short-range song (see Figure 2).

Figure 2 A free-living male dark-eyed junco courts a caged female during a behavioral trial. The male has erect body feathers and spread tail feathers, two visual displays associated with courtship behavior © Joe Welklin
We found that males that were presented with resident females exhibited more courtship behaviors than males that were presented with migrant females. When presented with resident females, males spent more time singing short-range song, the courtship song. Males also exhibited more visual displays when presented with a resident female: males puffed the body feathers and spread their tails more.
So, it appears that males can detect a potential resident versus migrant mate, suggesting that males may benefit from being able to discriminate females based on her reproductive timing. Based on this study, we cannot determine if males are preferring females based on their reproductive condition or other physical traits that distinguish residents from migrants. In either case, seasonal sympatry may facilitate the evolution of male mating preferences and these mating preferences may in turn contribute to maintaining population divergence.
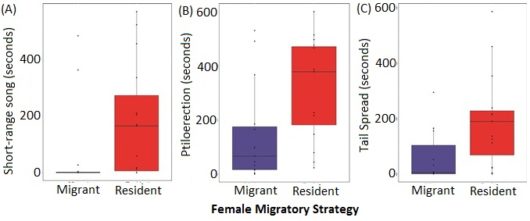
Figure 3 Males presented with resident females (A) sang more courtship song, (B) puffed their body feathers more, and (C) spread their tails more than males presented with migrant females
Now, as global change threatens biodiversity, we know that migratory birds are facing new challenges as they depend on multiple habitats over the course of an annual cycle. Both population ranges and timing of migration across many species are shifting in response to climate change. While this is just a small piece of the puzzle, understanding the mechanisms of divergence among populations that differ in migratory behavior may be important for understanding whether divergence will persist in the face changing climate. Scientists need to better understand the behavior and physiology of natural populations in order to determine the effects of climate change on biodiversity.
Nominate this article for a BOU Science Communication Award.
References
Ketterson, E.D., Fudickar, A.M., Atwell, J.W. & Greives, T.J. 2015. Seasonal timing and population divergence: when to breed, when to migrate. Current Opinion in Behavioral Sciences 6: 50-58. VIEW
Rolland, J., Jiguet, F., Jønsson, K.A., Condamine, F.L. & Morlon, H. 2014. Settling down of seasonal migrants promotes bird diversification. Proc. R. Soc. B 281: 20140473. VIEW
Williams, T.D. 2012. Physiological adaptations for breeding in birds. Princeton University Press, New Jersey, USA.
Winker, K. 2010. On the origin of species through heteropatric differentiation: a review and a model of speciation in migratory animals. Ornithological Monographs 69: 1-30. VIEW
Image credits
Featured image: Dark-eyed Junco (Oregon) | Becky Matsubara | CC BY 2.0

