

LINKED PAPER
Response of male Hawaii Elepaio Chasiempis sandwichensis to conspecific songs: A small-scale playback study. Ardeola. Oñate-Casado, J., Sebastián-González, E. & Hart, P.J. 2020. DOI: 10.13157/arla.67.2.2020.ra10. VIEW
In 2016, I got my first opportunity to collaborate in an international research group at the Hart Lab (University of Hawaii at Hilo). Everything started with an email to Dr Patrick Hart before the beginning of the last year of my undergrad. Since I started my university studies, I was keenly interested in ecology and animal behaviour. At that moment, I wanted to acquire bioacoustics research skills and link them to behavioural ecology. So when I came across a new bioacoustics lab making great strides in ecology, I did not hesitate to contact its head. A few emails and Skype calls with Dr Hart and his then post-doc Dr Esther Sebastián González, and I was completely convinced that this was a great opportunity. When I was in Hawaii a few months later, I did not know that this collaboration would result in my first first-author publication.
On the north-eastern slope of Mauna Loa Volcano there is an area of small fragments of undisturbed rainforest surrounded by young lava flows, called kipuka. Our study was conducted in two different sites within this area, both formed by several kipuka and separated by approximately 5 km (Fig. 1). I remember driving the lab´s pickup truck on the Saddle Road above Hilo, towards the foothills of the volcano. Once there, I used to walk off-road until one of the kipuka was reached. It is curious because at first sight, they seem to be simple small patches of vegetation, but once you enter it is just incredible how everything suddenly changes. In just a few seconds, you pass from walking across a “desert” of solidified lava to become surrounded by a dense rainforest that appears to have no end.


Figure 1 Map of Hawaii Island (left side) and the study area on the north-eastern slope of Mauna loa Volcano (right side). The white box indicates the study area. Kipuka sites are highlighted darker. The two white rectangular areas show one of the kipuka sites while the other site is represented by a single white-circled area. Individuals from three kipuka were considered at the low kipuka site while individuals from two kipuka were considered at the high kipuka site. The dashed line indicates the road near the low kipuka area
Kipuka are especially interesting because as fragmented areas, they are expected to have a reduced animal population size. This could decrease gene flow among individuals, leading to changes among fragments in certain social behaviours like acoustic traits (Laiolo & Tella 2005; Hart et al. 2018). But are changes in acoustic traits of these individuals accompanied by behavioural changes?
Kipuka host several species of endemic birds, among them, the Hawaii `Elepaio (Chasiempis sandwichensis; Fig 2). It is a monarch flycatcher endemic to the island of Hawaii in the Hawaiian archipelago (USA) and one of the most important birds in Hawaiian tradition, having several significant roles in culture and mythology. For instance, it is considered the ‘aumakua, or guardian spirit, of canoe makers: a physical manifestation of the canoe-building goddess, Lea. The `Elepaio also maintains year-round territories and is the least mobile of the species already investigated in this area. Its natural history coupled with its low density in the kipuka area made it the best option for our playback study.


Figure 2 Illustration of male (lower) and female Hawaii `Elepaio (Chasiempis sandwichensis) by Sailee Pradeep Sakhalkar
Previously, Sebastián-González & Hart (2017) observed an overall increase in acoustic dissimilarity with distance in nearby forest fragments for the Hawaii `Elepaio. This suggested that habitat fragmentation might produce cultural variation in song leading to dialect formation in neighbouring populations. In addition, it has been observed before that song variation may occur in Hawaii on a small scale, even from one valley to the next (e.g. through mist-netting `Elepaio using song playbacks, Eric VanderWerf, personal communication), but this had not been tested before our study.
We evaluated individual responsiveness to conspecific vocalisations within and between two nearby bird populations to assess within-species behavioural changes with distance. To do so, we used song playbacks of male ’Elepaio in a fragmented landscape. We then evaluated the response of males to songs recorded in their own populations and to those recorded in another population. Following the method of VanderWerf (2007), we measured individual response with three variables: vocal response, physical response, and latency. Vocal and physical response corresponded to the number of songs and the approaching distance to the speaker, respectively, while latency was the time interval between the stimulation (sound sequence) and response (vocal and/or physical). Furthermore, we also compared vocalisations between both populations to assess acoustic differentiation from recordings of wild male `Elepaio individuals taken previously from June 2014 to April 2015 by Sebastián-González and Hart (2017) in the same study area.
Our results indicate behavioural divergence in song responsiveness over a small spatial scale for this species (for latency but not for vocal nor physical response; Fig.3) and demonstrate that there is acoustic differentiation among `Elepaio vocalisations from nearby populations in a naturally fragmented landscape. However, contrary to what was expected, male individuals from both populations did not vary in their response pattern to songs depending on their origin. This suggests that acoustic differentiation may not be strong enough to drive behavioural divergence in the response of ’Elepaio individuals despite the existence of differences in the acoustic traits between both populations.
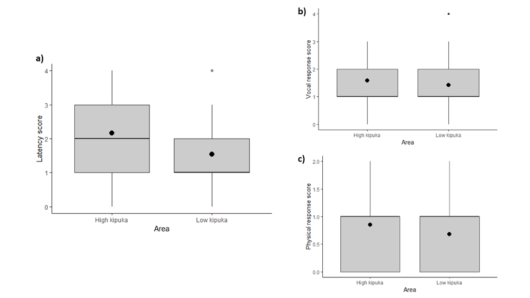
Figure 3 Boxplots of response scores (latency (a), vocal response (b), and physical response (c)) of subject ’Elepaio to playback songs of individuals from the same kipuka and a different kipuka. Black dots represent mean values, horizontal lines represent medians, vertical lines represent the upper and lower SE bars. Data are pooled across the low and high kipuka sites
This study shows that small-scale habitat fragmentation can affect song variation and elicit dialect formation by `Elepaio at this location. Fragmentation seems to act as a barrier that triggers acoustic variability, and consequently, might also affect the response to conspecific songs. Despite the results obtained, we need to continue to monitor more birds from this species across the range as we see them respond to their fragmented habitats.
References
Hart, P.J., Sebastián-González, S., Tanimoto, A., Thompson, A., Speetjens, T., Hopkins, M. & Atencio-Picado M. 2018. Birdsong characteristics are related to fragment size in a neotropical forest. Animal Behaviour 137: 45-42. VIEW
Laiolo, P. & Tella, J.L. 2005. Habitat fragmentation affects culture transmission patterns of song matchings in Dupont’s Lark. Journal of Applied Ecology 42: 118-193. VIEW
Sebastián-González, E. & Hart, P.J. 2017. Birdsong meme diversity in a habitat landscape depends on landscape and species characteristics. Oikos 126: 1511-1521. VIEW
VanderWerf, E.A. 2007. Bi0geography of ‘Elepaio: evidence from inter-island song playbacks. The Wilson Journal of Ornithology 119: 325-333. VIEW
Image credit
Featured image: Hawaii `Elepaio (Chasiempis sandwichensis) perched on a branch © Ann M. Tanimoto-Johnson

