

 LINKED PAPER
LINKED PAPER
Song varies with latitude, climate, and species richness in a Neotropical bird. Silva-Jr, E.F., Diniz, P. & Macedo, R.H. 2021 Behavioral Ecology. doi: 10.1093/beheco/arab112 VIEW
The world changes as we move across its boundaries. However, some extraordinary variations are not easily noted as we move from one place to another. A few years ago, I read a remarkable study on House Wren (Troglodytes aedon) song variation, and I remember being fascinated that this bird’s song complexity increased towards higher latitudes (i.e., the poles) in both northern and southern hemispheres across the Americas (Kaluthota et al. 2016). As you might imagine, a next step was asking: how does birdsong vary across other species’ distributions? Interestingly, two main sets of hypotheses are commonly used to clarify the pattern of increased birdsong complexity across broad geographic dimensions: (1) ecological hypotheses and (2) sexual selection hypotheses (Najar & Benedict 2019).
The ecological hypotheses predict increased song complexity as acoustic space availability becomes larger and/or vegetation properties become less sound-attenuating. For instance, where a bird coexists with more interfering sounds, such as other animals’ vocalizations and human noise, there is a lower frequency space availability, which can potentially constrain the evolution of complex songs (see Weir et al. 2012; Najar & Benedict 2019).
On the other hand, the sexual selection hypotheses predict a positive correlation between song complexity and sexual selection intensity. For example, imagine birds that inhabit high-seasonality localities (e.g., temperate regions at higher latitudes), where ideal amounts of resources to breed might be available only during a narrow window of time (Wyndham 1986). In such a condition, where birds have less time to breed – relative to other birds in low-seasonality localities –, sexual selection pressures might have been greater and have shaped sexual features that could reflect mate quality, like complex birdsongs, facilitating mate choice (see Catchpole 1982; Najar & Benedict 2019).
Overall, although birdsong complexity is a well-studied topic, there are few studies for large-scale regions, and the patterns of song complexity variation reported are not consistent (Najar & Benedict 2019). Considering this scenario, we determined patterns of song variation in the Blue-black Grassquit (Volatinia jacarina), a small bird distributed across the Neotropical region, more specifically from Mexico to Argentina (Rising 2020). To investigate the different hypotheses, we explored the relationship between the Grassquit song features, latitude, climate seasonality and bird species richness.
Blue-black Grassquit song
This bird species produces a monosyllabic song that is remarkably variable among individuals, even for those situated in the same locality (Fandiño-Mariño & Vielliard 2004).
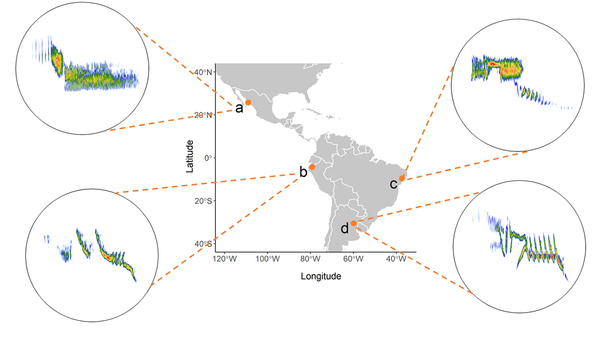
Figure 1 Spectrograms of Blue-black Grassquit songs recorded in distinct localities across the Neotropical range. Observe the remarkable variability in the structure of the song among different Grassquits. (a) Manuel Grosselet, (b) Jerome Fischer, (c) Pedro Têia, and (d) Bernabe Lopez-Lanus provided the recordings XC489743 (https://www.xeno-canto.org/489743), XC207196 (https://www.xeno-canto.org/207196), XC105631 (https://www.xeno-canto.org/105631), and XC47146 (https://www.xeno-canto.org/47146) to Xeno-Canto database, respectively.
To cover the species’ range, we gathered recordings from online databases of citizen science projects and audiovisual collections. We considered five acoustic metrics to indicate song complexity: entropy, frequency bandwidth, number of frequency inflections per second, number of song components and song duration (Benedict & Najar 2019; Crouch & Mason-Gamer 2019). Moreover, we calculated additional song metrics that could be under sexual selection pressures: median frequency, proportion of vibratos, and song rate (Linhart et al. 2012; Manica et al. 2014; Hill et al. 2015). Based on the hypotheses presented in the literature, we expected more complex songs at higher latitudes due to increased seasonality and species-poor habitats.
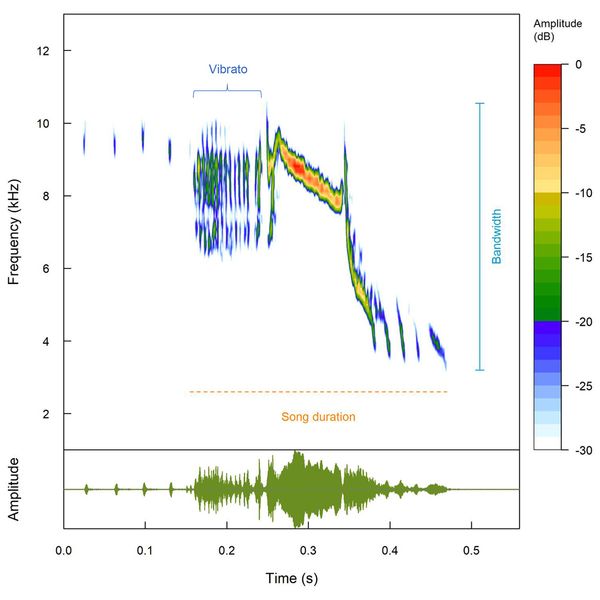
Figure 2 Spectrogram and oscillogram of a Blue-black Grassquit song with some acoustic features pointed out. Rodrigo Dela Rosa provided the recording XC70652 (https://www.xeno-canto.org/70652) to Xeno-Canto database.
Songs are mostly simpler at higher latitudes
Our findings are contrary to our primary expectation of more complex songs at higher latitudes. For the northern hemisphere, we observed that Blue-black Grassquit song contained fewer inflections per second and reached narrower frequency bandwidths as latitude increased. Additionally, for the southern hemisphere, song duration decreased as latitude increased. However, as expected, number of inflections correlated positively with latitude. Note that distinct song metrics varied in opposite directions within and between hemispheres, such as number of inflections and song duration in the southern hemisphere. Disparate results have also been indicated in other studies (e.g., Tietze et al. 2015), and such a fact undermines the explanatory power of ecological and sexual selection hypotheses regarding song complexity variation.
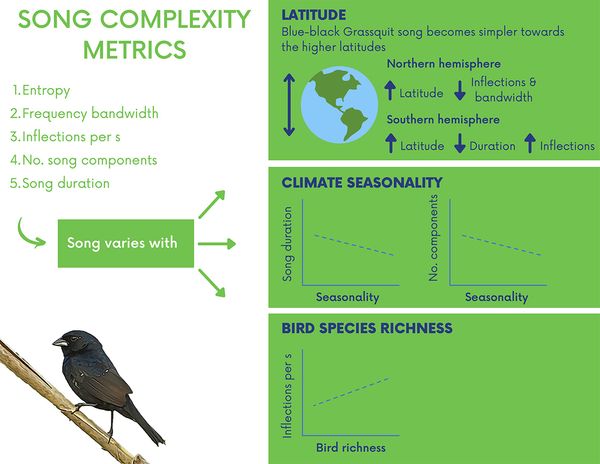
Figure 3 Acoustic metrics used to represent song complexity in the Blue-black Grassquit, and the main results of the study regarding the relationship between the Grassquit song complexity metrics, latitude, climate seasonality, and bird richness.
Simpler songs in more seasonal and species-poor environments
Based on sexual selection hypotheses, we expected more complex songs in habitats with greater climate seasonality (see Botero et al. 2009). Conversely, however, we found that song duration and number of song components of the Blue-black Grassquits decreased as climate seasonality increased. Moreover, contrary to our expectations based on ecological hypotheses, song complexity expressed by inflections per second correlated positively with bird species richness. In other words, Grassquits sing more complex songs where they coexist with more bird species potentially occupying the acoustic space available. Perhaps, in such a scenario, it could be beneficial to compress more acoustic information per unit of time. Lastly, we also observed that proportion of vibratos varied significantly with climate seasonality and may be a relevant song metric under sexual selection in the Grassquit.
Final thoughts
All in all, the Blue-black Grassquit song varies with latitude, climate and species richness. However, our results are mostly contrary to theoretical predictions based on ecological and sexual selection hypotheses. Taken together with results of other studies (Najar & Benedict 2019), we point out that these hypotheses do not fully explain song complexity variation for bird species distributed across broad geographic dimensions. Now, you could ask, what additional factors could affect birdsong variation across a species’ distribution? Future studies considering more species and other variables (e.g., historical isolation of populations, migration, and/or paleoclimate) will be necessary to clarify patterns of song evolution. Certainly, some extraordinary variations regarding birdsong still need to be uncovered.
An adapted version of this text is available in Portuguese on Edvaldo’s website.
References
Benedict, L. & Najar, N.A. 2019. Are commonly used metrics of bird song complexity concordant? The Auk 136: 1-11. VIEW
Botero, C.A., Boogert N.J., Vehrencamp, S.L. & Lovette, I.J. 2009. Climatic patterns predict the elaboration of song displays in mockingbirds. Current Biology 19: 1151-1155. VIEW
Catchpole, C.K. 1982. The evolution of bird sounds in relation to mating and spacing behavior. In D.E. Kroodsma & E.H. Miller (eds) Acoustic Communication in Birds: 297–319. New York, USA: Academic Press.
Crouch, N.M.A. & Mason-Gamer, R.J. 2019. Identifying ecological drivers of interspecific variation in song complexity in songbirds (Passeriformes, Passeri). Journal of Avian Biology 50: 1-14. VIEW
Fandiño-Mariño, H. & Vielliard, J.M.E. 2004. Complex communication signals: the case of the Blue-black grassquit Volatinia jacarina (Aves, Emberizidae) song. part I – a structural analysis. Anais da Academia Brasileira de Ciências 76: 325-334. VIEW
Hill, S.D., Amiot, C., Ludbrook, M.R. & Ji, W. 2015. Seasonal variation in the song structure of tui (Prosthemadera novaeseelandiae). New Zealand Journal of Ecology 39: 110-115. VIEW
Kaluthota, C., Brinkman, B.E., Dos Santos, E.B. & Rendall, D. 2016. Transcontinental latitudinal variation in song performance and complexity in house wrens (Troglodytes aedon). Proceedings of the Royal Society B 283: 1-8. VIEW
Linhart, P., Slabbekoorn, H. & Fuchs, R. 2012. The communicative significance of song frequency and song length in territorial chiffchaffs. Behavioral Ecology 23: 1338-1347. VIEW
Manica, L.T., Maia, R., Dias, A., Podos, J. & Macedo, R.H. 2014. Vocal output predicts territory quality in a Neotropical songbird. Behavioural Processes 109: 21-26. VIEW
Najar, N. & Benedict, L. 2019. The relationship between latitude, migration and the evolution of bird song complexity. IBIS 161: 1-12. VIEW
Rising, J.D. 2020. Blue-black Grassquit (Volatinia jacarina). In J. del Hoyo, A. Elliott, J. Sargatal, D. Christie & E. de Juana (eds) Birds of the World. New York, USA: Cornell Lab of Ornithology.
Tietze, D.T., Martens, J., Fischer, B.S., Sun, Y.H., Klussmann-Kolb, A. & Päckert, M. 2015. Evolution of leaf warbler songs (Aves: Phylloscopidae). Ecology and Evolution 5: 781-798. VIEW
Weir, J.T., Wheatcroft, D.J. & Price, T.D. 2012. The role of ecological constraint in driving the evolution of avian song frequency across a latitudinal gradient. Evolution 66: 2773-2783. VIEW
Wyndham, E. 1986. Length of birds’ breeding seasons. The American Naturalist 128: 155-164. VIEW
Image credit
Top right: Blue-black Grassquit Volatinia jacarina © Edvaldo Silva-Jr.
If you want to write about your research in #theBOUblog, then please see here.
