


A bird’s eye view of the Bay of Biscay
LINKED PAPER
Nocturnal bird migration in the Bay of Biscay as observed by a thermal-imaging camera. Weisshaupt, N., Maruri, M. & Arizaga, J. 2016. Bird Study. DOI: 10.1080/00063657.2016.1239693 VIEW
The East-Atlantic flyway represents one of the principal migration routes in Europe, comprising the Bay of Biscay as a geographical barrier for migratory landbirds. Few studies have been published concerning the role of the Bay of Biscay in bird migration. Broad-front migration was assumed based on diurnal visual and ringing data in autumn (Lack & Lack 1953). Arizaga et al. (2014) reported much weaker passage in spring compared to autumn at several coastal ringing sites along the Bay of Biscay. By contrast, a coastal operational radar located in the southeastern Bay of Biscay revealed high nocturnal activity in spring and low activity in autumn (Weisshaupt et al. 2014).
It is well known that variations in migration strategies found in short- and long-distance migrants cause changes in the spatial and seasonal flow of migration. Also prevailing seasonal meteorological conditions could have a favourable or deterrent effect on migrants’ motivation to travel. So basically, tailwinds (north- to north-easterly in autumn; south- to south-westerly in spring) both in autumn and spring could be expected to promote migration over the open sea of the Bay of Biscay. Some studies suggest, however, that birds also travel under suboptimal conditions, such as in headwinds or crosswinds, because waiting for optimal conditions would result in a major delay (Karlsson et al. 2011). Given the highly variable weather at the bay, it is possible thus that wind assistance plays a minor role.
Studying active nocturnal migration per se excludes certain sampling methods based on its very nature, such as ringing and visual counts. One available technique to study nocturnal activity is thermal-imaging (Zehnder et al. 2001).
One of the advantages is that thermal imaging allows us to obtain various migration parameters, such as migration intensity, direction and spatial arrangement of migrants (e.g. dense flocks vs. individuals). Flight altitudes can be estimated based on size classification of targets (Zehnder et al. 2001). A further benefit is the ease of use and quick start-up of such cameras. Their relatively small size and weight permit swapping easily between study sites within a short time. A disadvantage is that thermal- imaging cameras alone allows only for a rough discrimination between passerines and non-passerines based on individual flight (passerines) vs. flocking (non-passerines) (Zehnder et al. 2001).
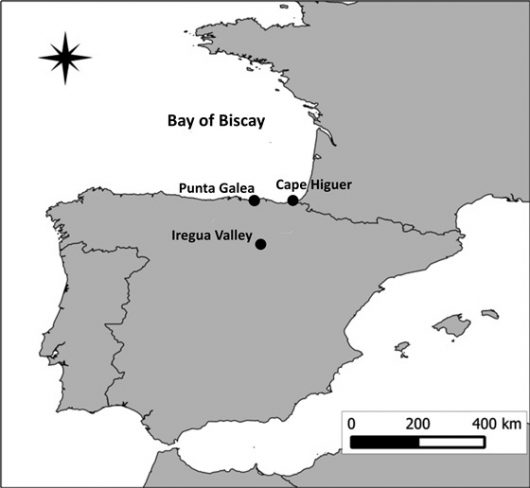 Figure 1 Locations of the three sampling sites used to quantify bird migration across the Bay of Biscay: Punta Galea, Cape Higuer and Iregua Valley (Northern Spain).
Figure 1 Locations of the three sampling sites used to quantify bird migration across the Bay of Biscay: Punta Galea, Cape Higuer and Iregua Valley (Northern Spain).
A thermal-imaging camera was used to study nocturnal bird migration patterns at two coastal study sites in Northern Spain: at Punta Galea (western coastal site) and Cape Higuer (eastern coastal site) in spring and at three study sites (coast: Punta Galea, Cape Higuer; inland: Iregua Valley) in autumn (Fig. 1). Sampling took place during four hours from sunset over 90 nights in 2014 and 2015 (Fig. 2).
 Figure 2 Thermal imaging camera set up at the meteorological station Punta Galea, Spain, at sunset
Figure 2 Thermal imaging camera set up at the meteorological station Punta Galea, Spain, at sunset
© Nadja Weisshaupt
Wind data at three height levels for the three sampling sites were collected from the National Oceanic and Atmospheric Administration, U.S. Department of Commerce, in order to check for a potential effect of wind directions and force on the observed migratory patterns.
Flight directions in spring pointed towards about 45°-60° at the coastal sites. In autumn, the both coastal sites showed more eastern directions (251°/255°) than the inland site (220°), indicating a directional shift towards south of migrants further inland. Reverse migration, i.e. migration in the opposite direction, occurred overall less frequently in spring (<2% of tracks) than in autumn (7.4–19.9% of tracks), which is probably due to the higher proportion of inexperienced juvenile birds in autumn.
Flight altitudes were generally high, with 80-100% of the tracks concentrating in estimated altitudes of about 2 km at all three sites and in both seasons. High flight altitudes have been generally linked with the proximity of ecological barriers.
Migration was strong at both coastal sites in March, decreasing towards April and May. Autumn migration was weak at the western coast, but strong at the eastern coast and inland. The majority of the targets were passerines, which could be deduced from the predominance of singly flying targets (Fig. 3a) over flocks (Fig. 3b). The peak in early spring suggests passage of short-distance migrants.
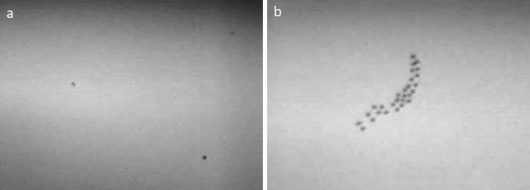 Figure 3 Two snapshots of nocturnal migration (a: passerines; b: non-passerines) at Punta Galea as seen by thermal imaging © Nadja Weisshaupt
Figure 3 Two snapshots of nocturnal migration (a: passerines; b: non-passerines) at Punta Galea as seen by thermal imaging © Nadja Weisshaupt
Tailwind did not have any significant effect on spatial and temporal variation of migration intensity. Despite that result, however, migration in spring ceased completely during strong easterly or northerly winds (i.e. direct or lateral headwinds) even though skies were clear. Thus, migrants may not show a clear preference for one particular wind direction, but simply for winds that would offer some tailwind component, and which may remain unappreciated in some of the analyses because of the wide range of wind directions.
Flight directions, altitudes, as well as seasonal and geographical variations in migration intensity, indicate that spring migration occurs on a broader front than autumn migration, potentially involving sea crossing. In autumn, there is a pronounced east–west gradient, indicating a more eastern migration flow.
Future studies should focus on the use of complementary long-range remote sensing technology, such as radar, to improve our understanding of migratory dynamics and patterns on a larger geographical scale at the Bay of Biscay.
Find out more about our research in the video below:
References and further reading
Arizaga, J., Fontanilles, P., Laso, M., Andueza, M., Unamuno, E., Azkona, A., Koenig, P. & Chauby, X. 2014. Stopover by reed-associated warblers Acrocephalus spp. in wetlands in the southeast of the Bay of Biscay during the autumn and spring passage. Revista Catalana d’Ornitología 30: 13–23. VIEW
Karlsson, H., Nilsson, C., Bäckman, J. & Alerstam, T. 2011. Nocturnal passerine migration without tailwind assistance. Ibis 153: 485–493. VIEW
Lack, D. & Lack, E. 1953. Visible migration through the Pyrenees: an autumn reconnaissance. Ibis 95: 271–309. VIEW
Weisshaupt, N., Maruri, M. & Arizaga, J. 2014. Bird migration across the Bay of Biscay observed by meteorological radar. Proceedings of ERAD 2014: 8th European conference on radar in meteorology and hydrology, Garmisch-Partenkirchen.
Weisshaupt, N., Maruri, M. & Arizaga, J. 2016. Preliminary assessment of nocturnal bird migration at the Bay of Biscay as observed by moon-watching. Munibe, 64. DOI: 10.21630/mcn.2016.64.03 VIEW
Zehnder, S., Åkesson, S., Liechti, F. & Bruderer, B. 2001. Nocturnal autumn bird migration at Falsterbo, South Sweden. J. Avian Biol. 32: 239–248. VIEW
Image credit
Featured image: Nocturnal migration © Nadja Weisshaupt
If you want to write about your research in #theBOUblog, then please see here.

