


LINKED PAPER
Nest attachment, rather than nest type, correlates with passerine bird brain size. Leite, A.B., Camacho, A., Francisco, M.R. 2024. IBIS. DOI: 10.1111/ibi.13292. VIEW
How do animals evolve to be smarter? Positive correlations have been found across different groups of organisms between residual brain size (relative to body mass) and the complexity of cognitively demanding activities, such as problem solving and social interactions. In birds, one of the most complex and likely cognitively demanding abilities is nest construction (Collias 1997), however, its relationship with brain size is not well understood. Among the vertebrates, birds have some of the largest brains relative to body size (Dunbar & Shultz 2007), but there is variation between species. Understanding the causes of this variation can help us understand how organisms with increased cognitive capacity evolved.
In a recent study in Ibis, Abraão Leite and colleagues used a dataset of 455 passerine bird species to address potential correlations between nest characteristics and relative brain mass.
The nest-building of passerines
Passerines are an ideal avian order for investigating the relationships between brain size and nest characteristics. Within Passeriformes, cup-shaped nests are the most common nest type, but this order also includes almost all bird families that construct enclosed nests, which have a roof and a lateral entrance (Martin et al. 2016). Passeriformes also represent the order in which nest attachment modes have reached the highest degree of diversification.
The researchers applied phylogenetic comparison methods to investigate the relationships between nest characteristics and passerine brain size by addressing both nest type (open and enclosed nests) and a proposed classification of 5 nest attachment modes (Basal A, Basal B, Lateral, Rim suspended, and Top suspended), controlled for body size, parental care, habitat type, and migratory tendencies. They addressed the premise that enclosed nests are more complex and require more cognitive abilities than open nests to construct, hence are associated with relatively larger brains. They also predicted that the building of nests held or suspended in vegetation by lateral or superior attachment points requires more complex weaving skills, and consequently larger brains, compared to nests positioned on flat surfaces.
Phylogenetic confirmatory path analysis was used to test several candidate predictive models to address whether nest components can interact with relative brain size via a direct evolutionary link, or via indirect links medicated through other variables, such as habitat. The evolutionary rates of brain size across nest component categories were also computed using a phylogenetic ridge regression approach to investigate whether the nest parameters were associated with brain size radiation or stabilisation.

Figure 1. Model-predicted relative brain mass (Rbmass) values across nest attachment categories organized in a proposed order of complexity. Coloured boxes represent the 25% quartiles, whiskers represent the 75% quartiles, horizontal lines represent the medians, asterisks represent the average values, points represent outliers and n is the number of species constructing nests of each attachment category.
Cognitive abilities are associated with nest attachment, not nest type
The results indicated that nest attachment mode, rather than nest type, was correlated with passerine relative brain sizes, despite the strong effects of habitat density, migration, and phylogeny. Relative brain mass (Rbmass) appeared to interact with nest attachment mode mainly in a direct way, consistent with a scenario in which a larger Rbmass is needed for the emergence of the most complex nest attachment modes. This model also supported an interaction to a lesser extent via the path mediated by habitat density. This could be due to the evolution of suspended nests being constrained by a lack of physical supports in certain habitats, for example the scarcity of trees and vines in habitats such as deserts or tundra. Further research using larger datasets could help to elucidate the relationships between nest attachment mode and habitat characteristics.
The results of the ridge regression analysis indicated that Top suspended nests were not associated with brain size diversification, but may have emerged in taxa already near a brain size maximum, consistent with the construction of nests with this attachment mode requiring more complex cognitive skills. The emergence of Rim suspended nests was found to be associated with passerine groups experiencing positive brain size radiations, which may indicate an interaction between brain size and Rim suspended nests in response to new ecological niches (Botton-Divet & Nyakatura 2021).
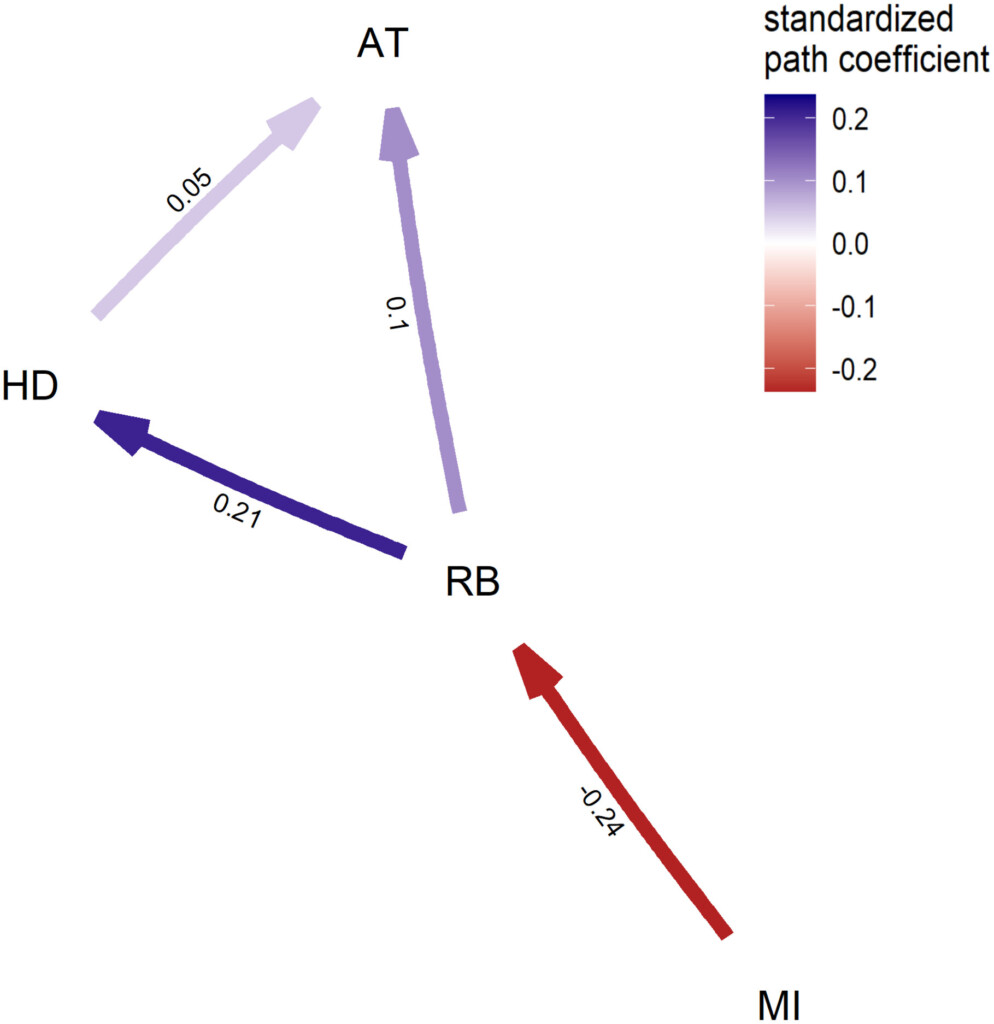
Figure 2. Best-fitting model from the confirmatory phylogenetic path analysis indicating that passerine relative brain mass (RB) interacts with nest attachment mode (AT) in a direct way, and to a lesser extent via a path mediated by habitat density (HD), with migration (MI) causing a negative direct effect on relative brain mass. Numbers associated with the arrows are the standardized correlation coefficients and colour intensity reflects the strength of positive (blue) or negative (red) interactions between a pair of variables.
While nest type was not found to be correlated with brain size in this study, it is possible that correlations may occur between nest type and the size or level of foliation of specific brain areas associated with cognitively demanding behaviours (Hall et al. 2013). However, the data necessary to investigate this is currently limited, meaning that Rbmass, as used in this study, remains the best available variable for assessing correlations between brain characteristics and avian cognitive capacity.
Overall, the results of this study indicate that the construction of nests with different attachment modes requires different levels of cognitive abilities, and the researchers provide insights into the understanding of the importance of brain size evolution for nest attachment diversification in passerines.
References
Botton-Divet, L. & Nyakatura, J.A. (2021). Vertical clinging and leaping induced evolutionary rate shifts in postcranial evolution of tamarins and marmosets (Primates, Callitrichidae). BMC Ecology and Evolution 21: 132. VIEW
Collias, N.E. (1997). On the origin and evolution of nest building by passerine birds. Condor 99: 253–270. VIEW
Dunbar, R.I.M. & Shultz, S. (2007). Evolution in the social brain. Science 317: 1344–1347. VIEW
Hall, Z.J., Street, S.E. & Healy, S.D. (2013). The evolution of cerebellum structure correlates with nest complexity. Biology Letters 9: 1–4. VIEW
Martin, T.E., Boyce, A.J., Fierro-Calderón, K., Mitchell, A.E., Armstad, C.E., Mouton, J.C. & Soudi, E.E.B. (2016). Enclosed nests may provide greater thermal than nest predation benefits compared with open nests across latitudes. Functional Ecology 31: 1231–1240. VIEW
Image credits
Top right: Four Curve-billed Thrasher eggs in a nest in Catalina Foothills, Tuscon, Arizona, USA | SearchNet Media | CC BY 2.0 Wikimedia Commons
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here

