


LINKED PAPER
Call rate as an index of nest count in wading bird colonies. Larson, R. C., Gawlik, D. E. 2023. IBIS. DOI: 10.1111/ibi.13163. VIEW
Across the globe, ecosystem degradation due to the pressures of climate change, pollution, and human activity is leading to the loss of biodiversity, habitats, and ecosystem services. While this ongoing crisis appears bleak, many projects to restore ecosystems are taking place to counteract the damage. However, a key factor in such programmes is detecting ecosystem changes and monitoring restoration progress, and this is often done using ecological indicators. While wading birds are a commonly used indicator of wetland health, the traditional methods (e.g. nest transects, aerial surveys, ground surveys) used to monitor them have numerous shortfalls.
In a recent study in Ibis, Rachel Larson and Dale Gawlik tested the use of acoustic monitoring as a colony monitoring technique for Snowy Egret (Egretta thula), Tricolored Heron (Egretta tricolour), and Great Egret (Ardea alba) colonies in south central Florida.
Monitoring wading bird colonies
Wading bird colony sizes and locations, timing of nesting, and productivity are metrics of wetland restoration progress as part of the Comprehensive Everglades Restoration Plan in South Florida, USA, which aims to restore the ecological function of the Everglades in response to anthropogenic disruptions in the natural hydropattern (RECOVER 2005). Traditional monitoring methods can be used to collect this data, but these can be disruptive to wildlife and it can be difficult to access wetland colonies to survey them, and methods such as aerial nest counts can be time-consuming and unreliable for detecting cryptic species (Rodgers et al. 2005).
Passive acoustic sampling is an increasingly common alternative method in such circumstances, where autonomous recording units (ARUs) are used to detect vocal activity. These require minimal set up, can be scheduled to record at specific times and dates, and can sample continuously across large spatial and temporal scales, providing flexibility and reducing the level of researcher disruption (Shonfield & Bayne 2017). The addition of signal recognition software can also enable automatic detection and classification of calls by species, increasing the efficiency of processing large acoustic datasets (Knight et al. 2017, Shonfield & Bayne 2017).
While previous studies have quantified the relationship between nest count or density and vocal activity, they have often focused on adult call rate (e.g. Borker et al. 2014). However, chick call rate may be a more accurate measure of nest count than adult call rate, since chicks are direct indicators of active nests. Hence, the researchers quantified the relationship between small heron (Snowy Egret and Tricoloured Heron), and Great Egret nest count and chick call rate in colonies located in a dynamic shallow lake system. They also evaluated the effects of environmental, physiological, and temporal factors on call detection, as acoustic detection can be influenced by soundscape variables.
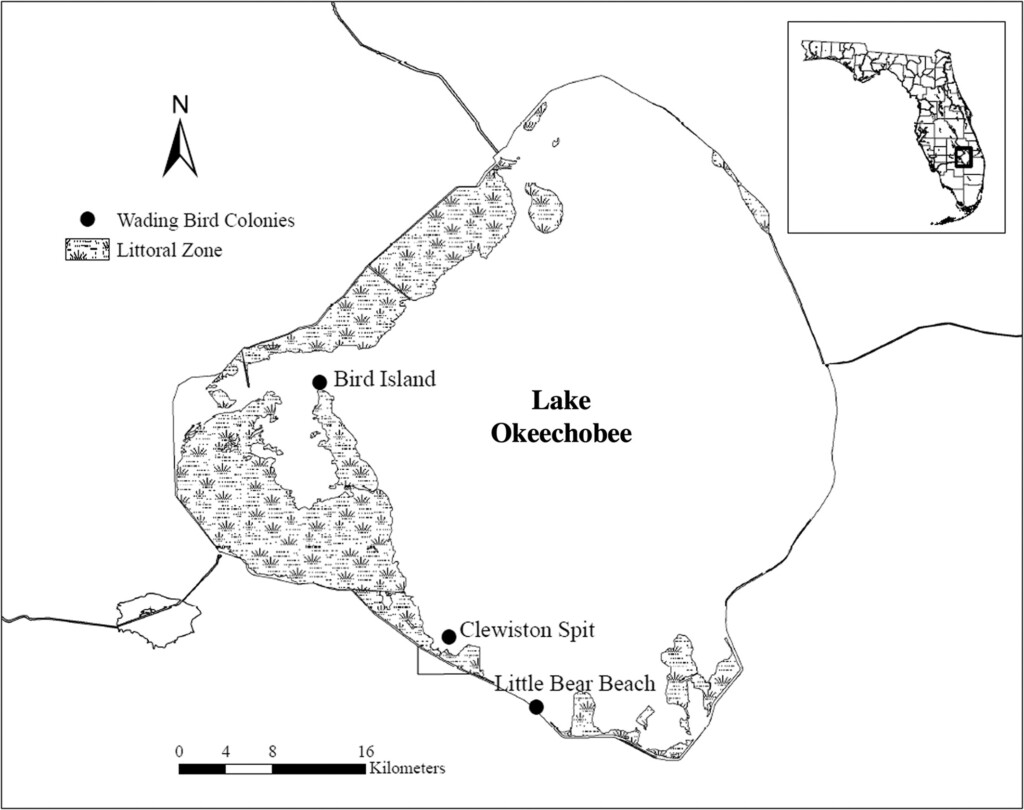
Figure 1. Researchers deployed autonomous recording units at Bird Island (56 small heron nests in 2019, 32 small heron nests in 2020) and Little Bear Beach (17 Great Egret nests in 2019, 22 Great Egret nests in 2020) located on Lake Okeechobee, Florida, USA.
The impact of external factors on acoustic methods
The results of generalised linear mixed-effects models showed that call rate detection increased with daily nest count for all species. While the detected call rate for small heron was dependent on all the measured variables, the Great Egret call detection decreased only with vegetation density around nests and daily wind speed.
Various factors such as weather conditions and habitat type have previously been shown to influence both acoustic methods (with detection by ARUs) and manual survey methods (with detection by humans) (e.g. Darras et al. 2018, 2019). For small herons, this study’s results indicated that call rate detection is influenced by day since colony initiation and body condition. It is possible that the age and physical condition of the chicks impacted the true call rate, resulting in the variation in detected call rate within the colonies. For all species, the detected call rates decreased with the density of vegetation between nests and the ARU and with the daily wind speed, which could be attributed to vegetation scattering and absorbing sound (Naguib & Wiley 2001), and to reduced detectability due to increased ambient noise (Buxton & Jones 2012).
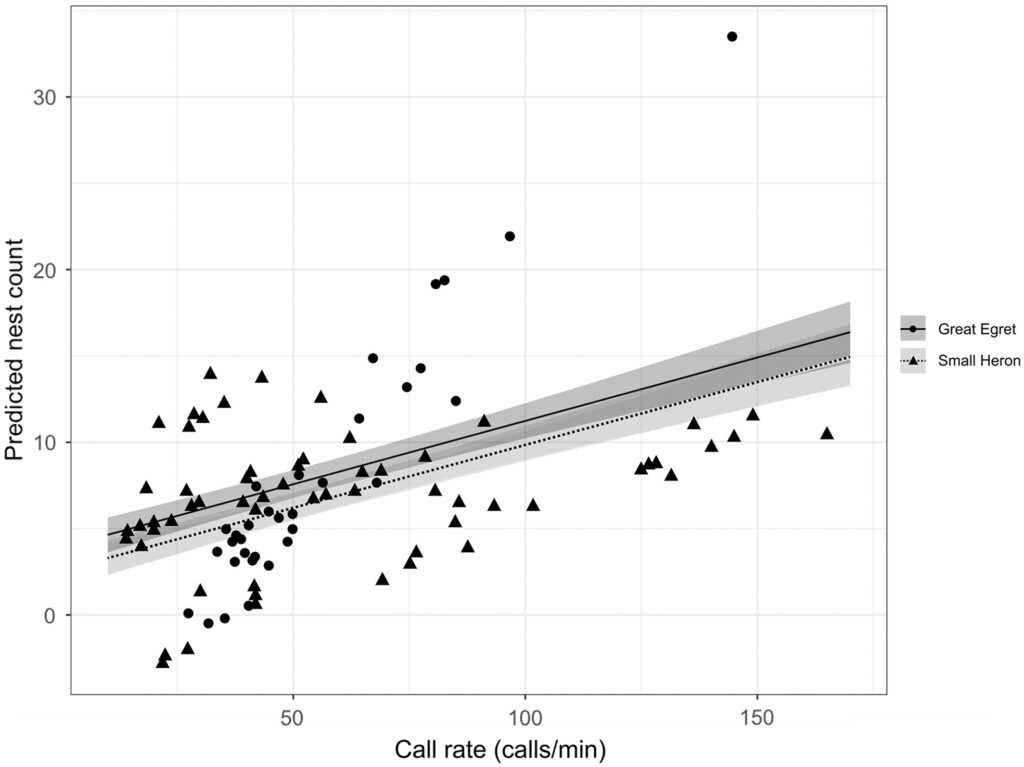
Figure 2. Predicted small heron and Great Egret nest counts derived from the top models explaining call rate. Small heron nest count is estimated from y = (β1 – 361 – 1.9β2 + 2.67β3 + 0.91β4 + 1.9β5 – 1.31β6)/(cs + 0.34β7) and Great Egret nest count is estimated from y.
The results demonstrated that passive acoustic sampling can be used successfully to estimate daily nest counts of small herons and Great Egrets throughout the breeding season. However, processing acoustic data was found to be time-consuming, and calls needed to be manually identified to develop a signal recogniser in the absence of a pre-existing one for heron chicks, and automatically classified calls also required manual verification. Further technological development, and advancements in machine learning software, are likely to improve data processing accuracy and efficiency in the future. This study also highlighted the importance of taking external factors into consideration during acoustic sampling, and including relevant variables in models.
References
Borker, A.L., Mckown, M.W., Ackerman, J.T., Eagles-Smith, C.A., Tershy, B.R. & Croll, D.A. (2014). Vocal activity as a low cost and scalable index of seabird colony size: Automated acoustic seabird monitoring. Conservation Biology 28: 1100–1108. VIEW
Buxton, R.T. & Jones, I.L. (2012). Measuring nocturnal seabird activity and status using acoustic recording devices: Applications for Island restoration. Journal of Field Ornithology 83: 47–60. VIEW
Darras, K., Batáry, P., Furnas, B., Celis-Murillo, A., Van Wilgenburg, S.L., Mulyani, Y.A. & Tscharntke, T. (2018). Comparing the sampling performance of sound recorders versus point counts in bird surveys: A meta-analysis. Journal of Applied Ecology 55: 2575–2586. VIEW
Darras, K., Batáry, P., Furnas, B.J., Grass, I., Mulyani, Y.A. & Tscharntke, T. (2019). Autonomous sound recording outperforms human observation for sampling birds: A systematic map and user guide. Ecological Applications 6: 1–19. VIEW
Knight, E.C., Hannah, K.C., Foley, G.J., Scott, C.D., Brigham, R.M. & Bayne, E. (2017). Recommendations for acoustic recognizer performance assessment with application to five common automated signal recognition programs. Avian Conservation and Ecology 12: 1–29. VIEW
Naguib, M. & Wiley, R. (2001). Estimating the distance to a source of sound: Mechanisms and adaptations for long-range communication. Animal Behaviour 62: 825–837. VIEW
RECOVER. (2005). The Recover Team’s recommendations for interim goals and interim targets for the Comprehensive Everglades restoration plan. Restoration Coordination and Verification Team (RECOVER), c/o United States Army Corps of Engineers, Jacksonville District, Jacksonville, Florida, and South Florida Water Management District, West Palm Beach, Florida.
Rodgers, J.A., Kubilis, P.S. & Nesbitt, S.A. (2005). Accuracy of aerial surveys of waterbird colonies. Waterbirds 28: 230–237. VIEW
Shonfield, J. & Bayne, E.M. (2017). Autonomous recording units in avian ecological research: Current use and future applications. Avian Conservation and Ecology 1: 1–13. VIEW
Image credits
Top right: Tricolored Heron – Egretta tricolor, Wakodahatchee Wetlands, Boynton Beach, Florida | Judy Gallagher | CC BY 2.0 Wikimedia Commons
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here

