


Is the Eurasian Magpie Pica pica better treated as five different species?
LINKED PAPER
Complete taxon sampling of the avian genus Pica (magpies) reveals ancient relictual populations and synchronous Late-Pleistocene demographic expansion across the Northern Hemisphere. Song, G., Zhang, R., Alström, P., Irestedt, M., Cai, T., Qu, Y., Ericson, P.G.P., Fjeldså, J. & Lei, F. 2018. Journal of Avian Biology. DOI: 10.1111/jav.01612. VIEW
The Eurasian Magpie Pica pica is widely distributed across the temperate parts of Eurasia, with isolated populations in Northwest Africa, Saudi Arabia and northeast Russia (Fig. 1). At least 10 subspecies are usually recognised. The Northwest African and Saudi Arabian subspecies have recently been suggested to be treated as two separate species, Maghreb Magpie Pica mauritanica and Asir Magpie Pica asirensis, respectively, based on plumage and structural characteristics (del Hoyo & Collar 2016). In addition, there are two species in North America, Black-billed Magpie Pica hudsonia and Yellow-billed Magpie Pica nutalli. The former was only rather recently split from Eurasian Magpie based on morphological, behavioural and mitochondrial restriction fragment profiles (Banks et al. 2000).
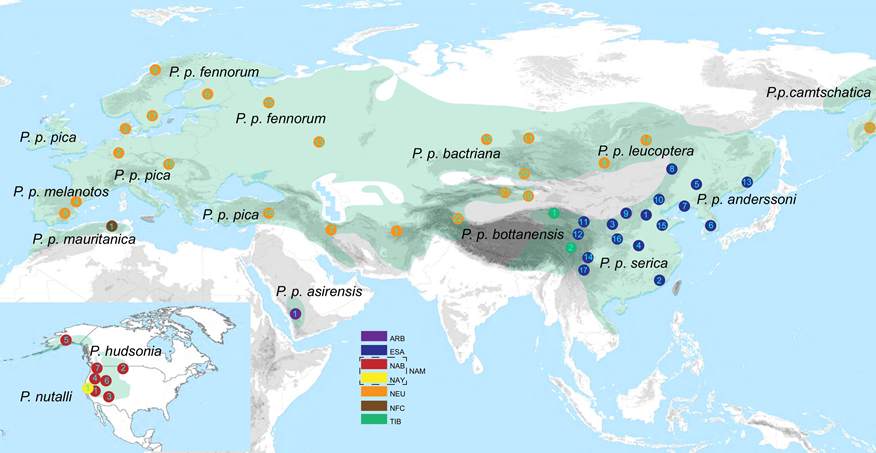 Figure 1 Distributions and sampling sites. Sampling sites are marked by coloured dots with numbers referring to locality names. Colours represent clades defined in Fig. 2. Click on image to view larger
Figure 1 Distributions and sampling sites. Sampling sites are marked by coloured dots with numbers referring to locality names. Colours represent clades defined in Fig. 2. Click on image to view larger
Six main groups, mainly separated during the Pleistocene
We analysed two mitochondrial and two nuclear DNA markers from all species and subspecies of Pica magpies. The mitochondrial tree identifies six main groups, which inhabit different geographical areas (Fig. 2). These are estimated to have diverged 1.4–3.1 million years ago.
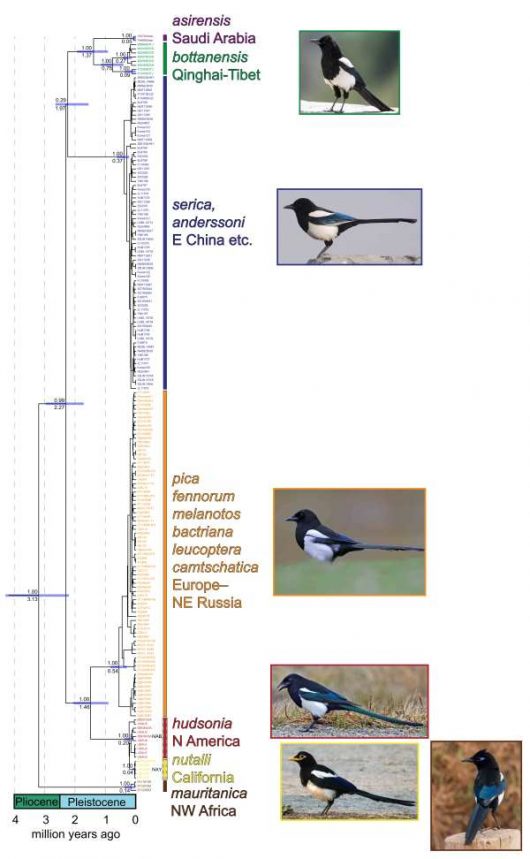 Figure 2 Mitochondrial DNA tree. Values above branches indicate Bayesian posterior probabilities (PP), and values below branches indicate the median value of the estimated age. Blue bars at nodes represent 95% highest posterior distribution (HPD). Nodes without values or 95% HPD bars are poorly supported (PP <0.90)
Figure 2 Mitochondrial DNA tree. Values above branches indicate Bayesian posterior probabilities (PP), and values below branches indicate the median value of the estimated age. Blue bars at nodes represent 95% highest posterior distribution (HPD). Nodes without values or 95% HPD bars are poorly supported (PP <0.90)
The Northwest African taxon, P. pica mauritanica, was the first one to become isolated from the others. All magpies from Europe to northeast Russia are genetically very similar. They are most closely related to the two North American species, which in turn are undifferentiated from each other in the analyzed markers. The Tibetan P. p. bottanensis and Saudi Arabian P. p. asirensis are suggested to be sister taxa, although they have been separated for approximately 1.4 million years. The two east Asian taxa P. p. serica and P. p. anderssoni are indistinguishable in the studied markers, but they are deeply diverged from the other groups since c. 2 million years.
Dispersal and isolation during climatic fluctuations
We suggest that magpies expanded their ranges during cooler periods of the Plio- and Pleistocene, when open grass- and scrublands were extensive, and that they later became isolated in smaller areas during periods of higher temperatures. Besides large water bodies (Mediterranean, Pacific), both extremely arid areas and dense montane forests may have constrained dispersal among populations.
For example, the peculiar distributional pattern of the sister pair P. p. bottanensis from the eastern Tibetan plateau and P. p. asirensis from southwestern Saudi Arabia is probably the result of fragmentation of a formerly continuous distribution of a common ancestor to these two, which became separated into two widely disjunct areas as a result of expansion of arid areas in intervening regions.
Magpies most probably colonized North America from eastern Asia. There is fossil evidence of range shifts within North America. Magpies were more widespread on that continent in ancient times, and were widely distributed in the southeastern part of the continent as late as the latest glaciation, at ca 12 000 yr ago (Emslie 1998).
Synchronous Late-Pleistocene demographic expansion across the Northern Hemisphere
Our analyses suggest demographic expansion in all the three regional clades (east Asia, North America, and north Eurasia) within the last 0.02–0.06 million years. The consistent expansion pattern indicates that magpies across the Eurasian continent and North America experienced a population growth at the same cooling transition time between the last interglacial and the last glacial maximum.
Yellow-billed Magpie stands out in appearance but not genetically
The North American Yellow-billed Magpie is the most unique-looking of all magpies thanks to its bright yellow bill and bare yellow facial skin (cf. Fig. 2), but it is undifferentiated from the North American Black-billed Magpie in the few genes analysed in this study. A possible explanation is that they have become separated from each other quite recently, but that their appearances have evolved at a much faster rate. Alternatively, the lack of divergence might be the result of past introgression. Further studies are required to try to distinguish these scenarios.
Taxonomic implications
We suggest that seven species should be recognised: Eurasian Magpie Pica pica sensu stricto (comprising six subspecies from Europe to northeast Russia); Maghreb Magpie P. mauritanica (Northwest Africa); Asir Magpie P. asirensis (soutwest Saudi Arabia); Black-rumped Magpie P. bottanensis (eastern Tibetan plateau); Oriental Magpie P. serica (east China and neighbouring areas); Black-billed Magpie P. hudsonia (northwestern North America); and Yellow-billed Magpie P. nutalli (California).
Acknowledgements
We are grateful to Arnoud van den Berg, Tomas Carlberg, Xiangyu Guan and Ming Wei for photos, and Yang Liu for help obtaining photos.
References
Banks, R.C., Cicero, C., Dunn, J.L., Kratter, A.W., Ouellet, H., Rasmussen, P.C., Remsen, J.V. Jr, Rising, J.A. & Stotz, D.F. 2000. Forty-second supplement to the American Ornithologists’ Union check-list of North American birds. Auk 117: 847–858. VIEW
del Hoyo, J. & Collar, N.J. 2016. HBW and BirdLife International Illustrated Checklist of the Birds of the World. Vol. 2: Passerines. Lynx Edicions and BirdLife International.
Emslie, S.D. 1998. Avian community, climate and sea-level changes in the Plio-Pleistocene of the Florida peninsula. Ornithol. Monogr. 50: 1–113. VIEW
Image credits
Featured image: Eurasian Magpie Pica pica pica, Sweden, 29 January 2018 © Tomas Carlberg
Fig. 2:
Black-rumped Magpie Pica bottanensis, Nangqian, Qinghai, China 20 July 2017 © Ming Wei
Oriental Magpie Pica serica, Beijing, China, 4 February 2016 © Xiangyu Guan
Eurasian Magpie Pica pica pica, Sweden, February 2018 © Tomas Carlberg
Black-billed Magpie P. hudsonia, North of Jasper, Alberta © Alan D. Wilson/ naturespicsonline.com/Wikimedia Commons
Yellow-billed Magpie Pica nutalli, California, 9 August 2016 © Per Alström
Maghreb Magpie Pica mauritanica, Agadir, Morocco, 4 November 2017 © Arnoud B. van den Berg
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here.

