


Geographic variation in phenotype and molecular markers within the Blue-black Grosbeak
LINKED PAPER
Congruence of phenotypic and genetic variation at the subspecific level in a Neotropical passerine. García, N. C., Barreira, A. S., Lavinia, P. D. & Tubaro, P. L. 2016. IBIS DOI: 10.1111/ibi.12386. VIEW
Many species show intra-specific geographical variation, which is extremely interesting for researchers as it can provide an insight into evolutionary and speciation processes. The subspecies concept is commonly used in ornithology to categorize this variation. Most subspecies were defined on the basis of geographical differences in traits such as body size, plumage colour, migration behaviour, or vocalizations. With the development of molecular techniques, the focus moved from phenotypic or behavioural characters to the analyses of variation in genetic markers. And with the increasing availability of molecular phylogenies it became evident that genotypic and phenotypic patterns of variation are not always congruent below the species level (and sometimes at the species level too).
These incongruities can arise due to a discordance of biological processes acting on each level, as genetic markers are assumed to evolve in a neutral or nearly neutral fashion (due to random drift) while phenotype is often under selective pressures (either natural or sexual, or both). But they could be the result of methodological artefacts as well. Most subspecies were described before the simplest statistical analyses, such as the t-test, were developed (Remsen Jr., 2005). A modern re-analysis of phenotypic variation would probably show that a significant proportion of the subspecies delineated are in need of revision. In several Neotropical bird species the number of subspecies is similar to that of genetic clades, although their geographic boundaries do not necessarily match. Some examples are the Lesser Woodcreeper (Xiphorhynchus fuscus, Cabanne et al. 2008), the Ochre-bellied Flycatcher (Mionectes oleaginous, Miller et al. 2008) or the Rufous-tailed Hummingbird (Amazilia tzacatl, Miller et al. 2011).
We analyzed the congruence of geographical variation between phenotype and genotype in a Neotropical passerine, the Blue-black Grosbeak (Cyanocompsa cyanoides). This bird is found from southern Mexico to Brazil and it is currently divided into four subspecies that differ on size and plumage color, being these differences subtle to the human eye.
Bryson and collaborators (2014) first reported the existence of three reciprocally monophyletic lineages within the species: one grouping members of C. c. concreta and C. c. toddi, a second one corresponding to C. c. cyanoides, and the third one corresponding to C. c. rothschildii. They found that C. c. rothschildii – the only taxon that occurs east of the Andes – has been evolving independently for more than 3 Ma and suggested that it should be considered a separate species. But no detailed phenotypic analysis was available to support such a proposal.
 Figure 1 Museum specimens (from the American Museum of Natural History, NY) representing the four Blue-black Grosbeak subspecies. From top to bottom, Cyanocompsa cyanoides concreta, C. c. toddi, C. c. cyanoides and C. C. rothschildii. © Natalia García
Figure 1 Museum specimens (from the American Museum of Natural History, NY) representing the four Blue-black Grosbeak subspecies. From top to bottom, Cyanocompsa cyanoides concreta, C. c. toddi, C. c. cyanoides and C. C. rothschildii. © Natalia García
We complemented these previous findings with new analyses to see if phenotypic variation among subspecies is congruent with genetic differentiation. We analyzed two mitochondrial genetic markers as well as morphological characters (body weight, wing length, beak size) and plumage coloration. We also considered variation in song, a behavioural character that is learned in Oscines and therefore could be considered to be more plastic than body size or coloration.
We found significant differentiation in genetic markers, body size, plumage coloration and also in song. On the one hand, when comparing subspecies, differences in these phenotypic and behavioural traits are much greater when the comparisons are made between C. c. rothschildii and the other subspecies, similar to what happens when assessing genetic markers. Thus, both phenotypic and genotypic evidence indicate that this subspecies is the most differentiated one supporting its consideration as a separate species, Cyanocompsa rothschildii.
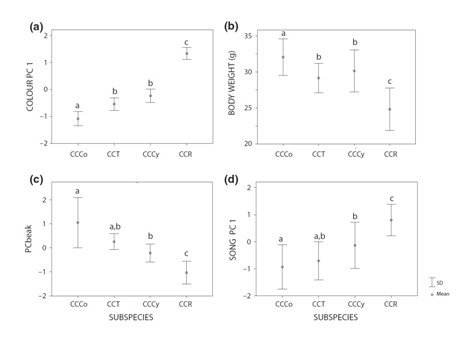 Figure 2 Variation between subspecies Cyanocompsa cyanoides concreta (CCCo), C. c. toddi (CCT), C. c. cyanoides (CCCy) and C. C. rothschildii (CCR) in plumage coloration (a), body weight (b), beak size (c) and song (d)
Figure 2 Variation between subspecies Cyanocompsa cyanoides concreta (CCCo), C. c. toddi (CCT), C. c. cyanoides (CCCy) and C. C. rothschildii (CCR) in plumage coloration (a), body weight (b), beak size (c) and song (d)
On the other hand, if phenotypic variation was fully congruent with genotypic differentiation, we would expect to find significant differences between C. c. cyanoides and the other two subspecies, while we would expect smaller or no differences between C. c. concreta and C. c. toddi. However, both body size and plumage coloration showed no significant differences between C. c. cyanoides and C. c. toddi, while these two differed significantly from C. c. concreta in both phenotypic traits. Interestingly, the trait that varied in a more similar manner to molecular markers was the song. Many aspects of song differed when comparing C. c. rothschildii with any of the other subspecies, only a few varied when comparing C. c. cyanoides to C. c. concreta and C. c. toddi, but none differed between these last two.
The partial congruence we found between phenotype and genetic markers shows that traditional classification of subspecies based solely on phenotype still carries valuable information on diversification and variation below the level of species. A great deal of criticism against the subspecies concept came after realization that many Nearctic and Palaearctic subspecies show null or reduced levels of genetic differentiation (Zink 2004). On the contrary, Neotropical species have shown in general greater levels of phylogeographic structure. In this context, our results also point out how the predominance of studies focusing on continental subspecies from North America and Eurasia, together with the gap of knowledge in Neotropical species, could bias our comprehension of general patterns of avian diversification.
References
Bryson, R. W. Jr, Chaves, J., Tilston Smith, B., Miller, M. J., Winker, K., Pérez-Emán, J. L. and Klicka, J. 2014. Diversification across the New World within the ‘blue’ cardinals (Aves: Cardinalidae). Journal of Biogeography. 41: 587-599. VIEW
Cabanne, G. S., d’Horta, F. M., Sari, E. H., Santos, F. R., & Miyaki, C. Y. 2008. Nuclear and mitochondrial phylogeography of the Atlantic forest endemic Xiphorhynchusfuscus (Aves: Dendrocolaptidae): Biogeography and systematics implications. Molecular phylogenetics and evolution. 49: 760-773. VIEW
Miller, M.J., Bermingham, E., Klicka, J., Escalante, P., do Amaral, F.S.R., Weir, J.T. & Winker, K. 2008. Out of Amazonia again and again: episodic crossing of the Andes promotes diversification in a lowland forest flycatcher. Proceedings of the Royal Society of London B: Biological Sciences. 275: 1133–1142. VIEW
Miller, M.J., Lelevier, M.J., Bermingham, E., Klicka, J.T., Escalante, P. & Winker, K. 2011. Phylogeography of the Rufous-tailed Hummingbird (Amazilia tzacatl). Condor. 113: 806–816. VIEW
Remsen Jr, J.V. 2005. Pattern, process, and rigor meet classification. Auk. 122:.403–413. VIEW
Zink, R. M. 2004. The role of subspecies in obscuring avian biological diversity and misleading conservation policy. Proceedings of the Royal Society of London B: Biological Sciences. 271: 561-564. VIEW
Further reading
Zamudio, K. R., Bell, R. C., & Mason, N. A. 2016. Phenotypes in phylogeography: Species’ traits, environmental variation, and vertebrate diversification. Proceedings of the National Academy of Sciences. 113: 8041-8048. VIEW
Image credit
Top Right: Blue-black Grosbeak Cyanocompsa cyanoides © Brian Gratwicke
If you want to write about your research in #theBOUblog, then please see here.

