

LINKED PAPER
Leg rings impact the diving performance of a foot-propelled diver. Gómez-Laich, A., Pantano, C., Wilson, R.P., Svagelj, W.S., Yoda, K., Gunner, R & Quintana, F. 2021 IBIS. doi: 10.1111/ibi.12998 VIEW
 Marking birds on their legs with aluminium or plastic bands with a unique combination of numbers, colours and/or letters, allows scientists to keep track of individuals whenever they might be caught or even seen again. Indeed, this approach has proved pivotal to many seminal studies examining, for example, bird dispersal, their migration and how long they live. The approach is also used for diving birds except penguins, where flipper, rather than leg bands were favoured until recently. Flipper bands, which are placed around the base of the wings (critical for generation of thrust to swim), can impair blood circulation during moult (Ainley et al. 1983). Critically, these bands also increase the costs of swimming (see Jackson & Wilson 2002) to the point where they can reduce penguin survival and breeding success (Saraux et al. 2011).
Marking birds on their legs with aluminium or plastic bands with a unique combination of numbers, colours and/or letters, allows scientists to keep track of individuals whenever they might be caught or even seen again. Indeed, this approach has proved pivotal to many seminal studies examining, for example, bird dispersal, their migration and how long they live. The approach is also used for diving birds except penguins, where flipper, rather than leg bands were favoured until recently. Flipper bands, which are placed around the base of the wings (critical for generation of thrust to swim), can impair blood circulation during moult (Ainley et al. 1983). Critically, these bands also increase the costs of swimming (see Jackson & Wilson 2002) to the point where they can reduce penguin survival and breeding success (Saraux et al. 2011).
Leg rings on foot-propelled birds
Findings from the penguin research got us thinking about the potential effects of rings on the legs of foot-propelled diving birds such as cormorants and grebes. Even though ringing diving birds on their legs is a common and worldwide practice, might it also affect diving behaviour and/or performance?
Over the past 15 years, several Imperial Cormorants (Leucocarbo atriceps) from the Punta León colony, Chubut, Argentina were ringed on both tarsi in order to facilitate behavioural studies. We used these birds to examine diving behaviour and compared them to individuals without leg rings. To do this, we equipped both groups with depth sensors and triaxial accelerometers allowing us to quantify the frequency and intensity of swimming movements as well as bird performance (Yoda et al. 1999). Although Imperial Cormorant dives have three clear distinct phases (a descent, a bottom phase and an ascent), our study focused only on the descent phase because at this time cormorants need to swim hard against their own buoyancy that tends to push them back up to the surface. Unsurprisingly, to overcome this buoyancy force birds kick more often and more vigorously than when they are at the seafloor searching for food or when ascending to the surface. Therefore, we suspected that ring effects, if any, would be most apparent during this phase of the dive.
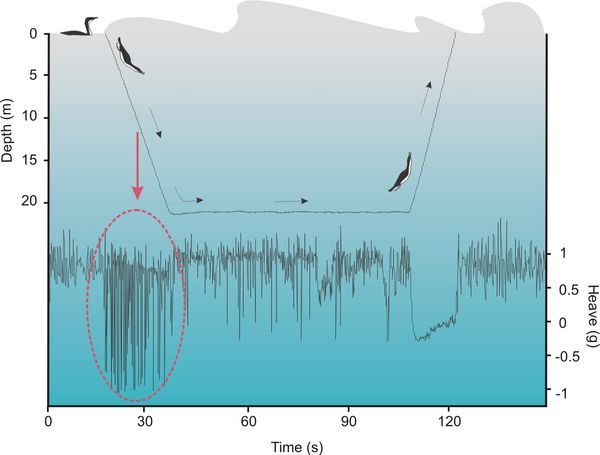 Figure 1 Example of the heave acceleration (up and down motion) signal from an accelerometer tag on an Imperial Cormorant (Leucocarbo atriceps) during the three distinct phases of a single dive. Note the clear peaks in the heave acceleration channel while descending along the water column to overcome the force of the bird buoyancy. These peaks represent foot kicks.
Figure 1 Example of the heave acceleration (up and down motion) signal from an accelerometer tag on an Imperial Cormorant (Leucocarbo atriceps) during the three distinct phases of a single dive. Note the clear peaks in the heave acceleration channel while descending along the water column to overcome the force of the bird buoyancy. These peaks represent foot kicks.
Kicking characteristics, energy expenditure and descent efficiency
As cormorants swam deeper during their descents, the frequency of foot strokes of both ringed and unringed birds decreased. This was because of the buoyancy force generated by the volume of air in the plumage decreasing with increasing pressure/depth. However, females with rings kicked more frequently than unringed birds at depths below 20 metres (m). Kick amplitude, an indicator of the strength of kicking, was also higher in ringed than unringed birds, principally above the first 20 m of the descent phase for females and below 20 m depth for males.
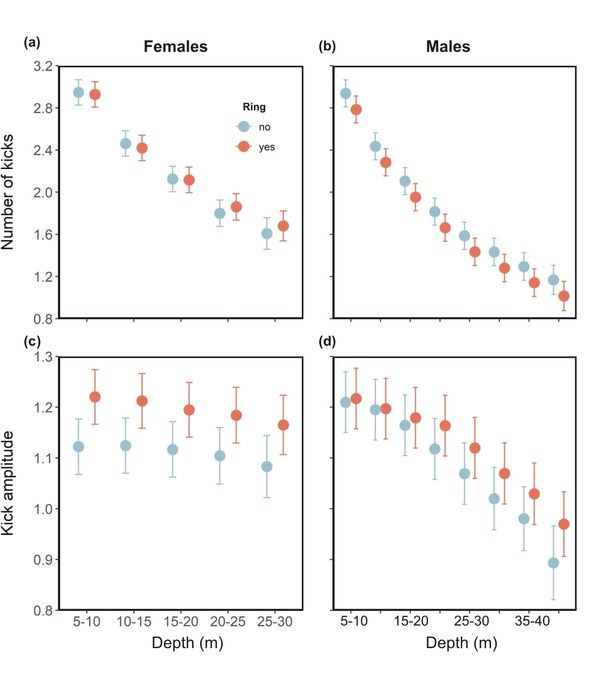 Figure 2 Male and female ringed Imperial Cormorants react differently to unringed birds: (a) Foot kick frequency and (b) kick amplitude in the various groups as a function of depth during dive descents.
Figure 2 Male and female ringed Imperial Cormorants react differently to unringed birds: (a) Foot kick frequency and (b) kick amplitude in the various groups as a function of depth during dive descents.
Using an acceleration metric known as VeDBA (Vectorial Dynamic Body Acceleration) (Qasem et al. 2012) as a proxy for energy expenditure, we found that female cormorants with rings worked harder to gain depths between 10 and 20 m than unringed birds. In males, VeDBA was similar between both ringed and unringed birds, except below 35 m where, curiously, VeDBA was higher for unringed birds. To be meaningful, kicking must be considered with respect to performance. Here, cormorants descended the water column at rates that ranged between 0.50 and 2.37 metres per second. However, females descended at similar rates irrespective of whether they were ringed or not until a depth of 15 m, after which ringed birds descended faster. Males without rings descended the water column faster than birds with rings and this difference was most obvious at depths between 5 and 20 m.
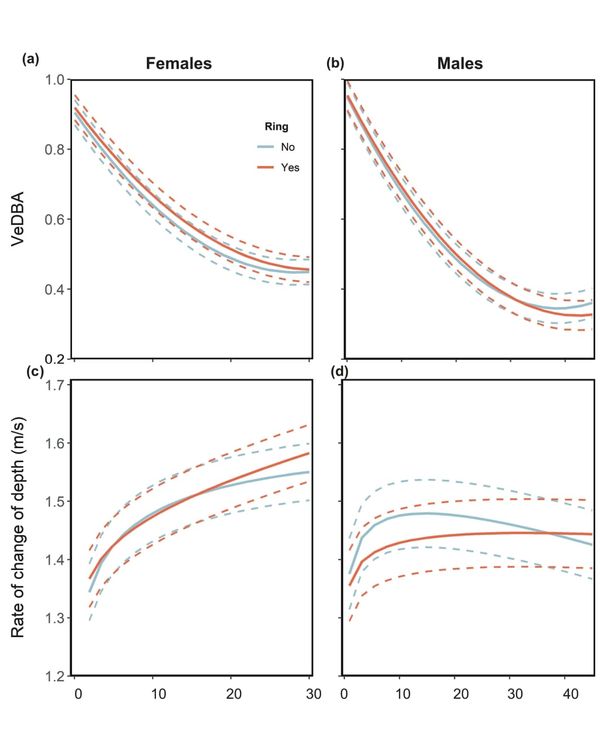
Figure 3 (a) and (b) VeDBA and (c) and (d) rate of change of depth in relation to depth during the descent phase of ringed and unringed female and male Imperial Cormorants. For both sexes, the continuous lines represent the fitted values obtained from a model while the dashed lines represent the 95% confidence intervals.
The crux of the story, however, lies in the birds’ efficiencies – how much effort is needed for a given swim performance for ringed and unringed birds? We examined this using the descent rate per unit of proxy for energy expenditure (VeDBA). This approach showed that our ringed birds worked harder for a given output than unringed birds, by about 3.5% for females and 4.3% for males across all depths.
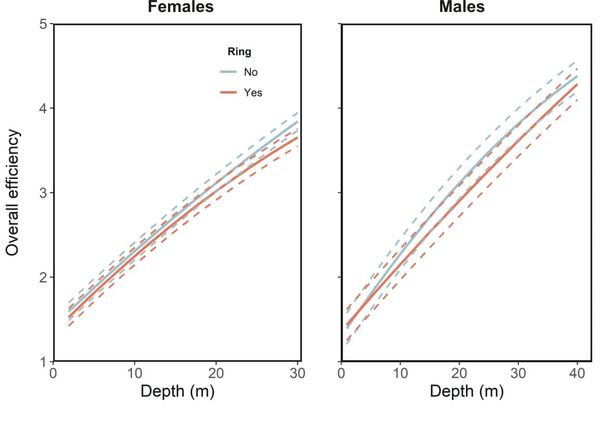
Figure 4 Overall efficiency (measured as the descent rate per unit VeDBA) in relation to depth for (a) female and (b) male Imperial Cormorants. For both sexes, the line represents the fitted values obtained from a model while the dashed lines represent the 95% confidence intervals.
Why would leg rings affect diving behaviour?
Being attached to the legs, which are used for propulsion in our birds, it is not difficult to conclude that rings may obstruct and decrease the efficiency of the propulsive stroke in a manner like that proposed for penguins. Although it is currently unclear precisely how rings might do this, future studies using techniques such as digital particle image velocimetry to examine flow patterns may provide an answer. Whatever the mechanism, the issue of decreased foraging efficiency due to the rings must be taken seriously, not least because such rings are effectively permanent meaning that deleterious effects, however small, may cause more substantial cumulative longer-term detriments to the behaviour of the bird. Although the small amount of detriment that we observed is unlikely to cause a reduction in breeding success, even in years of low prey availability, this work perhaps serves as a wake-up call to the general practice of ringing, in that we should always be vigilant to detriments to bird behaviour because of rings. In this, we should recognise that we are fortunate to work at a time when animal-attached technology can be used to elucidate important issues that cannot be resolved with the naked eye.
Image credit
Top right: Imperial shag (Leucocarbo atriceps) banded with a plastic ring © Agustina Gómez-Laich
References
Ainley, D.G., LeResche, R.E. & Sladen, W.J.L. 1983. Breeding biology of the Adélie penguin. University of California Press.
Jackson, S. & Wilson, R.P. 2002. The potential costs of flipper-bands to penguins. Functional Ecology 16: 141-148. VIEW
Qasem, L., Cardew, A., Wilson, A., Griffiths, I., Halsey, L.G., Shepard, E.L. & Wilson, R. 2012. Tri-axial dynamic acceleration as a proxy for animal energy expenditure; should we be summing values of calculating the vector? PLoS ONE 7: e31187. VIEW
Saraux, C., Le Bohec, C.L., Durant, J.M., Viblanc, V.A., Gauthier-Clerc, M., Beaune, D., Park, Y.H., Yoccoz, N.G., Stenseth, N.C. & Le Maho, Y. 2011. Reliability of flipper-banded penguins as indicators of climate change. Nature 469: 203-206. VIEW
Yoda, K., Sato, K., Niizuma, Y., Kurita, M., Bost, C., Le Maho, Y. & Naito, Y. 1999. Precise monitoring of porpoising behaviour of Adelie penguins determined using acceleration data loggers. Journal of Experimental Biology 202: 3121-3126. VIEW

