


Winter areas, spatial segregation and carry-over effects
LINKED PAPERS
Wintering areas predict age-related breeding phenology in a migratory passerine bird. López-Calderón, C., Hobson, K.A., Marzal, A., Balbontín, J., Reviriego, M., Magallanes, S., García-Longoria, L., de Lope, F. & Møller, A.P. 2017. Journal of Avian Biology. DOI: 10.1111/jav.01070. VIEW
Nowadays, an increasing number of papers are filling the knowledge gap about the winter ecology of migratory birds. This information is especially important in the context of global change, since many migratory birds are declining in population numbers and we do not even know where they spend half of the year. In addition, many studies recommend that aerial insectivores and long-distance migrants should receive special conservation concern (Sanderson et al. 2006, Nebel et al. 2010). Our study species, the House Martin Delichon urbicum, is one of these species, as its winter ecology is poorly known and its population trend is declining.
Very few House Martins have been ringed at their breeding areas and recaptured at their winter sites. The use of endogenous markers, such as stable isotope ratios in feathers, provides a powerful tool to determine winter origins of small passerines, which are not easy to track with external markers. In fact, the combination of ring recoveries and stable isotopes has been already used to infer the wintering areas of House Martins breeding in the Netherlands (Hobson et al. 2012a). Stable isotopes are useful not only for the identification of wintering areas, but also to determine a wide range of other winter ecology features.
In this study, we used stable isotope analyses (δ13C, δ15N, δ2H) to unravel three main characteristics of the winter ecology of House Martins breeding in southwestern Europe. Firstly, we aimed to identify their general wintering areas in Africa. Secondly, we asked if winter areas were spatially segregated according to age and sex. Thirdly, we tested whether winter habitat use had any carry-over effect in subsequent breeding phenology or reproductive success.
Our study took place in two breeding colonies of House Martins at Badajoz (38º52’N, 7º05’W), southern Spain, during 8 years (2005–2013, excluding 2006). Every year we captured more than 90% of the colony. Birds were sexed and categorized either as young (1 year old) or experienced (2 years or older). We ringed each bird, we measured tarsus and weight, and we removed the outermost rectrix for isotope analyses (original sample size of 300 birds). In one of our study colonies, we followed reproductive events to determine the start of laying and hatching.
We matched the previously described isotopic clusters proposed for Africa (Hobson et al. 2012b) with the isotopic proportions measured in our sample feathers. One third of the sampled House Martins were assigned to cluster 1 and two thirds to cluster 2. Both clusters were present in several main regions of Africa, thus we used a regression equation based on ring-recoveries to predict a wintering longitude of 0.5ºE. Based on this analysis, we used prior information on ring-recoveries to identify west Africa as the most-probable winter area (Fig. 1). Each isotopic cluster corresponded to a different African biome: moist broadleaf forest in “cluster 1” and savannah mixed with tropical seasonal forest in “cluster 2”.
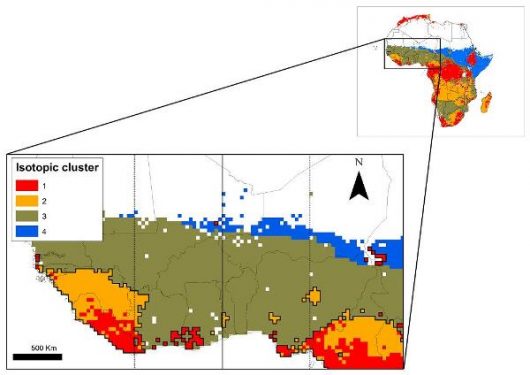 Figure 1 Predicted winter areas identified as isotopic clusters 1 and 2 throughout west Africa. The continuous and dotted vertical lines indicate the mean and 95% confidence intervals wintering longitude, estimated using linear regression analysis (Hobson et al. 2012a). Sample size is 300 sample feathers.
Figure 1 Predicted winter areas identified as isotopic clusters 1 and 2 throughout west Africa. The continuous and dotted vertical lines indicate the mean and 95% confidence intervals wintering longitude, estimated using linear regression analysis (Hobson et al. 2012a). Sample size is 300 sample feathers.
By means of linear mixed models, we detected that experienced males winter in isotopic cluster 2, with a higher probability than experienced females, and young females winter in the isotopic cluster 2, with a higher probability than young males (Fig. 2). After excluding the interaction term between age and sex, the linear mixed model confirmed that experienced House Martins winter in isotopic cluster 2 with a higher probability than young ones, regardless of sex.
 Figure 2 Relationship between age and sex on the probability to winter in isotopic cluster 2. Sample size is 45 experienced males, 8 young males, 39 experienced females and 15 young females wintering in isotopic cluster 2. Error bars indicate mean percentage ± 1 SE.
Figure 2 Relationship between age and sex on the probability to winter in isotopic cluster 2. Sample size is 45 experienced males, 8 young males, 39 experienced females and 15 young females wintering in isotopic cluster 2. Error bars indicate mean percentage ± 1 SE.
Furthermore, we found that young birds started to reproduce earlier than experienced ones when wintering in isotopic cluster 1 (one week on average), but there was no difference in breeding phenology of young and experienced birds birds wintering in isotopic cluster 2 (Fig. 3). After excluding the interaction term between age and isotopic cluster, the linear mixed model confirmed that birds wintering in isotopic cluster 2 bred earlier than birds wintering in isotopic cluster 1 (one week on average). By contrast, we did not find any difference in reproductive success of House Martins wintering in different areas.
 Figure 3 Relationship between age and isotopic cluster on breeding phenology. Black squares represent mean laying dates. Within the y-axis, “day 75” corresponds to 16 March and “day 150” to 30 May. Sample size is 112 birds.
Figure 3 Relationship between age and isotopic cluster on breeding phenology. Black squares represent mean laying dates. Within the y-axis, “day 75” corresponds to 16 March and “day 150” to 30 May. Sample size is 112 birds.
Previous studies have linked higher winter habitat quality to higher proportion of dominant individuals (Marra and Holmes 2001), earlier breeding phenology and improved reproductive success (Norris et al. 2004). We found that the proportion of experienced males is higher in isotopic cluster 2 of west Africa, and that birds wintering in isotopic cluster 2 reproduced earlier in the subsequent breeding season. Therefore, we propose that isotopic cluster 2 (tropical savannahs and seasonal forest) is of better quality winter habitat for House Martins than isotopic cluster 1 (broadleaf forest). We hypothesize that dominant House Martins (experienced males) would exclude subordinates (young males) from high quality winter habitats, reducing density in these habitats and finally experiencing a positive carry over effect in breeding phenology. Nevertheless, we acknowledge that fieldwork on the wintering areas is required to address these questions and so we need to get in touch with local researchers from west Africa.
In contrast to previous studies, we found that young birds wintering in isotopic cluster 1 lay their eggs earlier than experienced birds from the same wintering region. A plausible hypothesis to explain this result could be that experienced birds wintering in isotopic cluster 1 were of lower quality or senescent.
With this study, we have identified key features of the winter ecology of House Martins breeding in Spain, including wintering areas, spatial segregation and carry-over effects. However we have now several unsolved questions to continue working on until we know everything about the winter ecology of this interesting bird.
References and further reading
Hobson, K.A. et al. 2012a. Solving a migration riddle using isoscapes: House Martins from a Dutch village winter over West Africa. PLoS One 7: e45005. VIEW
Hobson, K.A. et al. 2012b. A multi-isotope (d13C, d15N, d2H) feather isoscape to assign Afrotropical migrant birds to origins. Ecosphere 3: 44. VIEW
Marra, P.P. & Holmes, R.T. 2001. Consequences of dominance-mediated habitat segregation in American Redstarts during the nonbreeding season. Auk 118: 92–104. VIEW
Nebel, S. et al. 2010. Declines of aerial insectivores in North America follow a geographic gradient. Avian Conserv. Ecol. 5: 1. VIEW
Norris, D.R. et al. 2004. Tropical winter habitat limits reproductive success on the temperate breeding grounds in a migratory bird. Proc. R. Soc. B-Biol. Sci. 271: 59–64. VIEW
Sanderson, F.J. et al. 2006. Long-term population declines in Afro-Palearctic migrant birds. Biol. Cons. 131: 93–105. VIEW
Image credit
Featured image: House Martin Delichon urbicum © Florentino de Lope
If you want to write about your research in #theBOUblog, then please see here.

