


Exploring bird associations across environmental and human disturbance gradients
LINKED PAPER
Measurement of species associations in mixed‐species bird flocks across environmental and human disturbance gradients. Mammides, C., Chen, J., Goodale, U.M., Kotagama, S.W., Goodale, E. 2018. Ecosphere. DOI: 10.1002/ecs2.2324. VIEW
Although we think of birds “of one feather flocking together”, actually many birds commonly interact in mixed-species flocks. These flocks can occur in migration and winter in temperate areas, and all-year round in the tropics, where in some forests as much as half of the birds are participating at any one time (Goodale et al. 2017). Watching these flocks can be one of the most exciting experiences a birdwatcher can have: in China for example, these flocks can have more than ten species and fifty individuals moving actively inside an area the size of a basketball court. This exciting phenomenon is also a great model for understanding principles of community organization. Further, it is not just theoretically interesting, for one fifth of the world’s species participate in these flocks, including many endangered species, and those species that are most important to the organization of flocks – known as “nuclear species” – can be targeted in conservation plans (Zou et al. 2018).
 Figure 1 View from the canopy top at one of the study areas, the Sinharaja Forest Reserve in Sri Lanka © Salindra Kasun Dayananda
Figure 1 View from the canopy top at one of the study areas, the Sinharaja Forest Reserve in Sri Lanka © Salindra Kasun Dayananda
In this study, we explored what influences the associations of species in flocks. Specifically, we had three questions:
First, associations between participating species have been measured using different methods but it is not known to what extent these methods are comparable. For example, traditionally the associations between species were measured using coefficients based on simple co-occurrences tables, such as the phi coefficient (Yule, 1912). Lately though, researchers have been assessing associations using tools derived from the social-networks methods (Whitehead, 2008; Farine et al. 2012). Additionally, a new approach has been applied (Srinivasan et al. 2010), which is based on simulated flocks. With the “null simulations” method, the associations are measured by comparing the observed flocks to flocks simulated according to each species’ total abundance (i.e., the number of individuals recorded inside and outside the flocks). Considering the variation in the methods, it was essential to evaluate whether the different approaches produce analogous results when applied to the same dataset.
Second, it is not known how consistent associations are across environmental and human disturbance gradients. For example, it has not been examined previously whether species that tend to associate strongly in one altitudinal level or land-use type (e.g., in low elevation or primary forest) are also consistently found together in another (e.g., in high elevation or farmland). This question of consistency is particularly important for understanding whether management of nuclear species can conserve multiple species as mentioned above, since this approach would only be efficient if a nuclear species’ role is consistent over a wide range of areas and habitats.
Third, it is unclear which species traits influence most the strength of the associations. Previous studies, have found that species with similar traits (e.g., in terms of body mass and diet) and congeneric species are more likely to associate with each other. Yet, the extent to which these results can be generalized needed to be explored.
To answer these questions, we used a well-replicated dataset of 221 bird flocks, collected at 41 transects in four areas in southwestern Sri Lanka. The study areas covered a wide range of altitudes (90 to 2180 m) and multiple land-use types (from primary forest in nature reserves to buffer zones and farmlands outside the reserves).
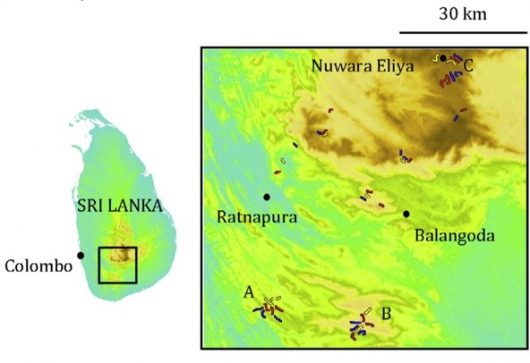 Figure 2 Map of the study areas and transects in Sri Lanka (adapted from Fig. 1 in Goodale et al. 2014)
Figure 2 Map of the study areas and transects in Sri Lanka (adapted from Fig. 1 in Goodale et al. 2014)
We found that indeed results varied depending on the method used. The pairwise correlations between the three methods ranged from 0.53 to 0.75. Specifically, the null simulations, which by default use also the out-of-flock bird abundances, correlated less with the other two methods: the phi-coefficient and the social networks, which traditionally have been applied using only the in-flock-data. However, once the out-of-flock abundances were added to these two methods too (by considering each single species and monospecific group as a separate flock), the results converged and the measured associations became more similar (0.75 < r < 0.94).
Regarding the second question, we found that associations across altitudinal and human-disturbance gradients varied substantially and only a small percentage of those associations were statistically significant. Although the exact values differed depending on the method used (and whether the out-of-flock data were added to the phi coefficient and social-networks methods), overall, the correlations between the associations at the various altitudinal levels and land-use types were low (r < 0.50). This suggests that the associations were not consistent but rather fluid. In other words, species that preferred to associate with each other at one altitudinal level or land-use type did not necessarily do so in another. We should note however that in this project we only had a few species considered nuclear, so we were unable to test whether nuclear species consistently have stronger associations with other species.
Finally, we found that on average similarity in body mass and diet resulted in stronger associations but the explanatory power of these two factors was low, regardless of the method used to measure the associations. The same was true for phylogenetic relatedness, for which contrary to a global meta-analysis (Sridhar et al. 2012), we found that congeneric species are less likely to associate with each other in our region.
Two important messages result from this work:
- It is apparent that the method used to measure the strength of the associations does influence the results, at least to a certain extent, and therefore researchers need to be aware of this. The same applies to when selecting whether to include or exclude the out-of-flock data when using the phi coefficient and the social-networks methods.
- Mixed-species bird flocks, at least in our study region, appear to be a fairly fluid system, in which on average the identity of the participating species does not determine which other species will join the flock or not. We stress “on average”: this does not take into account whether species play particularly important roles in flocks.
Overall, conservationists now view the interactions of species as important to conserve, just as much as the species themselves (e.g., Valiente-Banuet et al. 2015). We hope our research increases our understanding of how associations between species are measured, and how human disturbance influences them.
References
Farine, D.R., Garroway, C.J. & Sheldon, B.C. 2012. Social network analysis of mixed-species flocks: exploring the structure and evolution of interspecific social behaviour. Animal Behaviour 84(5): 1271-1277. VIEW
Goodale, E., Kotagama, S.W., Raman, T.S., Sidhu, S., Goodale, U., Parker, S., & Chen, J. 2014. The response of birds and mixed-species bird flocks to human-modified landscapes in Sri Lanka and southern India. Forest Ecology and Management 329: 384-392. VIEW
Goodale, E., Beauchamp, G. & Ruxton, G.D. 2017. Mixed-species Groups of Animals: Behavior, Community Structure, and Conservation. Academic Press, USA. VIEW
Sridhar, H., et al. 2012. Positive relationships between association strength and phenotypic similarity characterize the assembly of mixed-species bird flocks worldwide. American Naturalist 180: 777–790. VIEW
Srinivasan, U., Raza, R.H. & Quader, S. 2010. The nuclear question: rethinking species importance in multi-species animal groups. Journal of Animal Ecology 79: 948–954. VIEW
Valiente‐Banuet, A. et al. 2015. Beyond species loss: the extinction of ecological interactions in a changing world. Functional Ecology 29(3): 299-307. VIEW
Whitehead, H. 2008. Analyzing animal societies. University of Chicago Press, Chicago, Illinois, USA. VIEW
Yule, G.U. 1912. On the methods of measuring association between two attributes. Journal of Royal Statistical Society 75: 579–652. VIEW
Zou, F., Jones, H., Jiang, D., Lee, T.M., Martínez, A., Sieving, K., Zhang, M., Zhang, Q. & Goodale, E. 2018. The conservation implications of mixed-species flocking in terrestrial birds, a globally-distributed species interaction network. Biological Conservation 224: 267-276. VIEW
Image credit
Featured image: Sri Lanka Blue Magpie, Urocissa ornata © Salindra Kasun Dayananda
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here.

