


Can seabirds tell the difference between live and museum specimens?
LINKED PAPER
Feather colours of live birds and museum specimens look similar when viewed by seabirds.
Martin, A.B., Gaskett, A.C. & Friesen, M.R. 2017. IBIS. DOI: 10.1111/ibi.12501. VIEW
There are lots of reasons to envy birds: you’re almost always gorgeous if you’re a male and you can fly, usually, very gracefully. But there are even better reasons out there. In the last decade or so, we’re finally beginning to understand what the world looks like through a bird’s eyes, and it’s astonishing.
Humans have three types of colour receptor cells in our eyes, called cone cells, for detecting red, blue and green. Birds, however, have four (Chen & Goldsmith, 1986). Their fourth cone lets them see in the ultraviolet part of the spectrum, and it turns out that most birds that might have looked pretty uninteresting to us can actually glow when viewed in the right light.
For example, people once thought the Eurasian blue tit, Cyanistes caeruleus, was sexually monochromatic – males and females look the same colour to us. However, when researchers measured the spectra of light absorbed and reflected by the birds with a spectrophotometer, they found that blue tit plumage reflects ultraviolet light, and that it is sexually dichromatic, with males reflecting more UV from their head crests (Hun et al. 1998). UV bright male ornaments have also been found in the Australasian gannet, Morus serrator (Ismar, et al. 2014) king penguins (Jouventin et al, 2005). These UV patches might function in mate choice, much like the showy tails of Birds of Paradise (Pruett-Jones & Pruett-Jones, 1990) – so now we have a much greater insight into the behaviour of these birds.
We’re learning that birds have more colour receptors than us and that their plumage reflects a wider range of colours than we can see. How can we tell how what they see?
A basic understanding of species’ colour perception can be obtained by measuring the spectral reflectance of their plumage and skin using a spectrometer. Spectrometers can measure wavelengths of light both within and beyond the human visible spectrum (Eaton & Lanyon, 2003). Then, we can estimate bird perception of colour with models that incorporate the types and relative abundances of UV, blue, green and red photoreceptors, their sensitivities to different light wavelengths, and how the spectra reflected by an item excite those photoreceptors (Vorobyev & Osorio, 1998).
This sounds great, but how it is often difficult to use a spectrometer on live animals or in the field because the lens must be held still and very close to the colour patch … and every birder knows that birds can be hard to find and catch, can live in remote places, or may even be already extinct! Handling birds can also be stressful … to both bird and birder. To resolve this, many comparative studies of colour (comparing genders, or between geographic locations) are carried out on museum specimens (e.g. Bleiweiss, 2006 or Badyaev et al. 2002).
But to human eyes museum specimen’s colour can change over time, potentially compromising their usefulness in these endeavours (e.g. Doucet & Hill, 2009 or Daviset al. 2013). But very few have looked at this from a bird’s perspective. And nobody had looked at it from a seabird perspective. Can seabirds tell the difference between the colours of a live bird and a museum specimen?
Seabirds aren’t well known for their colours, but there are a few famous examples. In blue-footed Boobys, Sula nebouxii, male foot colour is bright blue. This bright blue colour varies depending on male quality: the brighter the better, and females will change their reproductive investment based on the foot colour of their partners (Velando, Beamonte-Barrientos, & Torres, 2006).
The seabirds commonly seen in New Zealand, Procellariiforms, are typically grey and white. The reasons behind this colour scheme is unclear – perhaps it is because they are nocturnal animals on breeding sites, and so don’t use colour vision that much. A recent study suggested that black and white colouration of seabirds exists to promote lift: the dark upper wing surface can be approximately 10 degrees Celsius warmer than the pale underside, and the temperature difference lowers the air pressure on the upperside of the wing – reducing drag and providing additional lift. This makes flight cheaper for birds, in terms of energy (Hassanalian et al. 2017).
Though their plumage may be boring, they do have different coloured feet: black, yellow, pink and in fifteen of the 125 species of Procelariiform, blue.
To figure out whether the colours of museum specimens are good proxies for live birds, we measured spectral data from live and stuffed birds of four species: Pelagodroma marina (White faced storm petrel), Pelecanoides urinatrix (Common Diving Petrel), Puffinus assimilis (Little Shearwater) and Puffinus gavia (Fluttering Shearwater).
Plumage or skin colour was measured on four patches: crown of the head, the top of the middle toe on the right foot, the dorsal feathers of the right wing, and the side of the bill inferior to the nares, with three measurements taken per body region. We used a package in R, a statistical programming software, called PAVO. We tested whether seabirds would perceive any differences in brightness and colourThe model calculates colour differences, in units aptly called, “Just Noticeable Differences” (JND). If the JND is 1 or greater, the bird can distinguish between two item’s colours. The larger the JND, the greater the colour difference.
We found that several seabirds had hidden UV reflectance – especially those birds with blue feet.
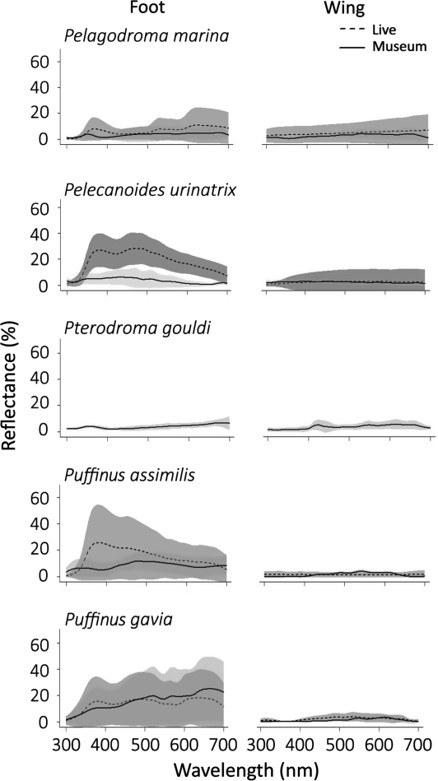
Figure 1 Mean ±sd (shaded) spectral reflectance of live and museum specimens of five seabird species
Seabirds would detect a difference between blue feet of museum and live birds, but luckily, not for the plumage.
 Figure 2 Pairwise comparison of live birds with museum specimens of varying age groups for the four species that had colour measurements for live and museum specimens. The scale on the right refers to JND: a black colour indicates a JND greater than or equal to 1, and as such are above the threshold of detectability of a difference for the seabird. Bird images courtesy of Aviceda (Pelagodroma marina), J. J. Harrison (Pelecanoides urinatrix), Terry Greene (Puffinis assimilis), Alan Tennyson (Puffinus gavia) and New Zealand Birds Online.
Figure 2 Pairwise comparison of live birds with museum specimens of varying age groups for the four species that had colour measurements for live and museum specimens. The scale on the right refers to JND: a black colour indicates a JND greater than or equal to 1, and as such are above the threshold of detectability of a difference for the seabird. Bird images courtesy of Aviceda (Pelagodroma marina), J. J. Harrison (Pelecanoides urinatrix), Terry Greene (Puffinis assimilis), Alan Tennyson (Puffinus gavia) and New Zealand Birds Online.
This means Museum specimen seabirds are excellent proxies for comparing plumage colours of live birds – saving birds and birders time, energy and stress! However, if you need to know about birds blue feet colours, you’ll still need to find and measure the real live bird to really get a bird’s eye view.
References
Badyaev, A.V., Hill, G.E., & Weckworth, B.V. 2002. Species divergence in sexually selected traits: increase in song elaboration is related to decrease in plumage ornamentation in finches. Evolution 56(2): 412-419. VIEW
Bleiweiss, R. 2006. Variation in ultraviolet reflectance by carotenoid-bearing feathers of tanagers (Thraupini: Emberizinae: Passeriformes). Biol. J. Linnean Soc. 84(2): 243-257. VIEW
Chen, D., & Goldsmith, T. 1986. Four spectral classes of cone in the retinas of birds. J. Comp. Physiol. 159(4): 473 – 479. VIEW
Davis, A.K., Woodall, N., Moskowitz, J.P., Castleberry, N., & Freeman, B.J. 2013. Temporal Change in Fur Color in Museum Specimens of Mammals: Reddish-Brown Species Get Redder with Storage Time. International Journal of Zoology 2013(2013): 876347. VIEW
Doucet, S.M., & Hill, G.E. 2009. Do museum specimens accurately represent wild birds? A case study of carotenoid, melanin, and structural colours in long-tailed manakins Chiroxiphia linearis. J. Avian. Biol. 40(2): 146-156. VIEW
Eaton, M.D., & Lanyon, S.M. 2003. The ubiquity of avian ultraviolet plumage reflectance. Proc. Biol. Sci. 270(1525): 1721-1726. VIEW
Hassanalian, M., Abdelmoula, H., Ayed, B., & Abdelkefi, A. 2017. Thermal impact of migrating birds’ wing color on their flight performance: Possibility of new generation of biologically inspired drones. Journal of Thermal Biology 66: 27-32. VIEW
Hunt, S., Bennett, A.T., Cuthill, I.C., & Griffiths, R. 1998. Blue tits are ultraviolet tits. Proc. Biol. Sci. 265(1395): 451 – 455. VIEW
Ismar, S.M.H., Daniel, C., Igic, B., Morrison-Whittle, P.K., Ballard, G., Millar, C.D., Fidler, A.E., McGraw, K.J., Wakamatsu, K., Stephenson, B.M., Cassey, P., Dearborn, D.C. & Hauber, M.E. 2014. Sexual plumage dichromatism in a size monomorphic seabird. The Wilson Journal of Ornithology 126(3): 417-428. VIEW
Jouventin, P., Nolan, P., Örnborg, J., & F Stephen, D. 2005. Ultraviolet beak spots in king and emperor penguins. The Condor 107(1): 144-150. VIEW doi:10.1650/7512
Pruett-Jones, S.G., & Pruett-Jones, M.A. 1990. Sexual selection through female choice in Lawes’ Parotia a lek-mating bird of paradise. Evolution 44(3): 486-501. VIEW doi:10.1111/j.1558-5646.1990.tb05934.x
Velando, A., Beamonte-Barrientos, R., & Torres, R. 2006. Pigment-based skin colour in the blue-footed booby: an honest signal of current condition used by females to adjust reproductive investment. Oecologia 149(3): 535-542. VIEW doi:10.1007/s00442-006-0457-5
Vorobyev, M., & Osorio, D. 1998. Receptor noise as a determinant of colour thresholds. Proc. Biol. Sci. 265(1394): 351-358. VIEW doi:10.1098/rspb.1998.0302
Image credit
Featured image: Little Shearwater, Puffinus assimilis © NZ Birds Online/Albert Aanensen
If you want to write about your research in #theBOUblog, then please see here.

