

LINKED PAPER
Habitat-use patterns among migrant and resident landbirds of contrasting dietary habits in a southern Mexican wetland. Ruiz, M.D.M., León-Cortés, J.L., Enríquez, P.L., García-Estrada, C. & Rangel Salazar, J.-L. 2019. Ardeola. DOI: 10.13157/arla.66.2.2019.ra3. VIEW
In general, research on migratory bird species at their temperate breeding grounds is relatively well consolidated when compared to research from the tropical latitudes where they overwinter. Nonetheless, a growing literature surrounds the use of coastal wetlands by Neotropical migratory landbirds (Fig 1). In Mexico, the coastal lagoons are scattered along 10,000 km of coastline, where they constitute perennially moist environments that provide resources for birds during the dry and rainy seasons.
Figure 1 Eight locations in the northern Neotropical region where assemblages of landbirds have been researched in coastal wetlands. Numbers correspond to studies in: 1) South-west Mexico (Ruiz et al., 2017); 2) south-east Mexico (Lynch, 1989); 3) Belize (Gómez-Montes & Bayly, 2010); 4) Jamaica (Johnson & Sherry, 2001); 5) Puerto Rico (Reitsma et al., 2002); 6) Costa Rica (Barrantes, 1998); 7) Panama (Lefebvre & Poulin, 1997); and 8) Venezuela (Lefebvre et al., 1994). In the inset, contour lines show 600 (black) and 1000 (grey) m a.s.l.
Each year many millions of landbirds seasonally relocate to Mexico (Robertson et al., 2011), yet how this staggering abundance is distributed across ecosystems is far from clear. In addition to their importance for migrant survival, Neotropical wintering grounds are also essential to an exceptionally rich resident avifauna. In this paper, we revealed guild-level habitat-use patterns among resident and migrant landbird species by analysing 32 consecutive months of standardized point count samples recorded at mangrove and marsh habitat types of a coastal lagoon in the south-western state of Oaxaca.
We disclosed temporal variation in local trophic guilds of insectivores and plant-consumers by examining monthly fluctuations (Fig 2). This initial appraisal showed that some migrants were present during every month of the year and highlighted their heavy reliance on the non-breeding distribution area. In particular, a relatively stable abundance of migratory insectivores from September to April pointed to the consistent use of mangroves.
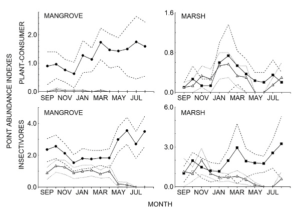
Figure 2 Monthly point abundance indices of resident (filled symbols) and migratory (triangles) birds from four local trophic guilds in the Ventanilla-Tonameca wetland, Oaxaca, Mexico, using data from October 2009 to May 2012. Bootstrap confidence intervals (0.95) are represented by dashed lines for resident birds and dotted lines for migratory, and the scale of the y-axes differs between graphs
We then used non-breeding season densities and average species abundances to quantify and compare the occurrence of resident and migratory birds in mangrove and marsh and evaluate the relative importance of these habitat types. Results showed that insectivorous migratory birds comprised over one third of their trophic guild density in both mangrove and marsh. For plant-consumers the mangrove held relatively few migrants, but in marsh they comprised almost half of the observed density.
With the notable exception of plant-consumers in mangrove, the average abundances of migratory and resident species in trophic guilds were comparable. These findings support the view that during their time in the Neotropics migrants are not the ephemeral species they were once considered to be, but are in fact integral members occupying stable niches in tropical ecosystems (Faaborg et al., 2010).
We evaluated two hypotheses that potentially explained our observed habitat-use patterns. Because they hinged upon the consumption of specific trophic resources, we focused on species subsets nested within trophic guilds: 14 species of frugivore (plant-consumers that mainly eat fruit) and 22 species of invertebrate gleaner (insectivores that mainly eat terrestrial leaf and stem invertebrates: Ruiz et al., 2017).
First, we developed a resource-centred dietary requirement hypothesis: We proposed that non-breeding season abundance among migrant frugivores in mangrove would be low, relative to marsh, because of the low availability of trophic resources for adult frugivores: mangrove trees produce fruits that are unpalatable and they are also devoid of any understorey that could produce alternative fruits. Conversely, we maintained that conditions in the mangrove would beneficial for resident frugivores, which require a high availability of potential nest-sites in order to reduce the risk of nest predation (Martin, 1993), and a relatively stable supply of a protein–rich invertebrates with which to provision their nestlings (Stutchbury & Morton, 2001).
Our analyses revealed lower migrant abundance in mangrove (average species abundance = 2) than in marsh (average species abundance = 32.5) and vice versa for resident species (average resident species abundance was 19.4 and 2.3 in mangrove and mash, respectively). A resident species held the highest abundance rank in mangrove (Yellow-winged Cacique Cassiculus melanicterus), whereas the counterpart in marsh was migratory (Orchard Oriole Icterus spurius Figure 3). This pattern was congruent with our resource-centred hypothesis of habitat-use by frugivores and the pattern was also evident in the wider trophic guild of plant-consumers.

Figure 3 The migratory Orchard Oriole Icterus spurius was more abundant than any other frugivore species at the marsh, with an estimated 1.7 ± 1.2 individuals per hectare © Mateo Ruiz
As an alternative proposition, we considered the nest predation hypothesis, which has been discussed as an explanation for habitat-use patterns among migrant warblers, and other species that are mainly insectivorous (Johnson et al., 2006). Using the assumptions of comparable trophic resource availability across habitat types and asymmetrical competition between resident and migratory species, this hypothesis predicts higher proportions of migrants in disturbed habitat types because limiting nest site availability should reduce competition for food with resident species. Accordingly, we predicted a greater non-breeding season abundance of resident invertebrate gleaners in the structurally complex mangroves than in non-woody marsh vegetation, which grows in areas with high flood plain disturbance.
However, our data did not uphold this prediction. For invertebrate gleaners the proportions of migrants in mangrove (0.45) and marsh (0.55) were comparable, and average species abundances were not markedly different (using a negative binomial GLM, Z = 0.95 and P = 0.34 for between-habitat type difference in resident species, and Z = 0.41 and P = 0.68 for migrants). We therefore concluded that any between-habitat variation in invertebrate gleaner nesting activity had not markedly influence non-breeding season abundance of resident or migratory species (Fig 4).
In this paper, we showed that where migratory and resident species assemble and how they interact can be contingent upon habitat type and guild membership. We maintain that resources in the coastal lagoons on the Mexican Pacific are used by an important abundance of both migratory and resident landbirds, as well as diverse waterbird assemblages (Mellink & de la Riva, 2005). Generalising habitat-use from species to guilds shed light on how the requirements of migratory and resident birds can drive assemblage-level patterns. In the future, we hope that research will be able to track and understand the seasonal movements of landbirds between coastal lagoons and surrounding habitat types.

Figure 4 A Sclater’s Wren Campylorhynchus humilis investigating invertebrate prey. We recorded a very similar abundance of this common resident species in both mangrove and marsh. Until recently the Sclater’s Wren was considered conspecific with the Rufus-naped Wren C. rufinucha © Mateo Ruiz
Nominate this article for a BOU Science Communication Award.
References
Barrantes, G. 1998. Reproductive activity of birds in a mangrove swamp in Northwest Costa Rica. International Journal of Tropical Biology and Conservation 46(4): 1163-1166. VIEW
Faaborg, J., Holmes, R.T., Anders, A.D., Bildstein, K.L., Dugger, K.M., Gauthreaux Jr., S.A., Heglund, P., Hobson, K.A., Jahn, A.E., Johnson, D.H., Latta, S.C., Levey, D.J., Marra, P.P., Merkord, C.L., Nol, E., Rothstein, S.I., Sherry, T.W., Sillett, T.S., Thompson III, F.R. & Warnock, N. 2010. Recent advances in understanding migration systems of New World land birds. Ecological Monographs 80(1): 3-48. VIEW
Gómez-Montes, C. & Bayly, N.J. 2010. Habitat use, abundance, and persistence of Neotropical migrant birds in a habitat matrix in northeast Belize. Journal of Field Ornithology 81(3): 237-251. VIEW
Johnson, M.D. & Sherry, T.W. 2001. Effects of food availability on the distribution of migratory warblers among habitats in Jamaica. Journal of Animal Ecology 70(4): 546-560. VIEW
Johnson, M.D., Strong, A.M. & Sherry, T.W. 2006. Migrants in tropical bird communities: the balanced breeding limitation hypothesis. Journal of Avian Biology 37(3): 229-237. VIEW
Lefebvre, G., Poulin, B. & McNeil, R. 1994. Temporal dynamics of mangrove bird communities in Venezuela with special reference to migrant warblers. Auk 111(2): 405-415. VIEW
Lefebvre, G. & Poulin, B. 1997. Bird communities in Panamanian black mangroves: potential effects of physical and biotic factors. Journal of Tropical Ecology 13(1): 97-113. VIEW
Lynch, J.F. 1989. Distribution of overwintering nearctic migrants in the Yucatan Peninsula, I: General patterns of occurrence. Condor 91(3): 515-544. VIEW
Martin, T.E. 1993. Nest predation and nest sites. BioScience 43(8): 523-532. VIEW
Mellink, E. & de la Riva, G. 2005. Non-breeding waterbirds at Laguna de Cuyutlán and its associated wetlands, Colima, Mexico (Aves acuáticas no-reproductivas en la laguna de Cuyutlán y sus anegados asociados en Colima, México). Journal of field ornithology 76(2): 158-167. VIEW
Reitsma, L., Hunt, P., Burson III, S.L. & Steele, B.B. 2002. Site fidelity and ephemeral habitat occupancy: Northern Waterthrush use of Puerto Rican black mangroves during the nonbreeding season. Wilson Bulletin 114(1): 99-105. VIEW
Robertson, B.A., MacDonald, R., Blancher, P.J. & Bevier, L. 2011. Boreal migrants in winter bird communities. In Wells, J.V. (ed.) Boreal Birds of North America, pp. 85-94. University of California Press. VIEW
Ruiz Bruce Taylor, M.D.M., Rangel Salazar, J.L., Enríquez, P.L., León-Cortés, J.L. & García-Estrada, C. 2017. Variation in hierarchical guild structure between two bird assemblages of a wetland in the Mexican Pacific. International Journal of Tropical Biology and Conservation 65(4): 1540-1553. VIEW
Stutchbury, B.J. & Morton, E.S. 2001. Behavioral Ecology of Tropical Birds. Academic Press, Bodmin, England. 165 p. VIEW
Image credits
Featured image: American Yellow Warbler Setophaga petechia © Matthew Twiggs

