


LINKED PAPER
Variability of a consistent trait: The size of the white wing patch in European Stonechats (Saxicola rubicola rubicola). Oficialdegui, F. J. & Serrano, D. 2024. IBIS. DOI: 10.1111/ibi.13252. VIEW
Sexual selection has been widely studied since it was first proposed by Charles Darwin in the 1800s. In short, as part of mate choice the “choosy” sex has a preference for mates displaying traits, e.g. ornaments, that signal either direct benefits (which increase the choosing partner’s fitness or survival), or indirect benefits (which give genetic advantages to offspring). A key assumption of this is that the traits must be honest signals of quality which are costly to produce and maintain, meaning they are limited to individuals in good condition. This also means that signals such as ornaments can vary over an individual’s life due to both genetic and environmental factors, as well as variables such as age.
Avian plumage ornaments have been reported as signals of individual quality in several different species. Differences between individuals in their striking white plumage patches have been well studied, but phenotypic variation and the factors affecting their consistency within individuals are less well known. In a recent study in Ibis, Francisco Oficialdegui and David Serrano studied a Spanish population of European Stonechats (Saxicola rubicola rubicola) over 6 years to analyse the variation in the size of the white wing patch, and identify its inter- and intra-annual dynamics.
European Stonechat plumage
While both male and female European Stonechats have white plumage patches, they exhibit pronounced sexual dimorphism. Adult males have prominent white patches in their wings, neck-sides, and rump, while females and juveniles are duller, with a cryptic coloration, apparently smaller white patches in their wings and neck-sides, and no white patch in the rump (Svensson et al. 1999). The species develop a new sexual ornament every year, with a partial moult from fledgling to first-year age, and a complete post-breeding moult afterwards. Linear mixed models (LMMs) were used to determine the correlates of wing patch size, with data from 431 individuals (220 males, 211 females) in total, and variables such as age, sex, individual condition, time from moult, and environmental variables.
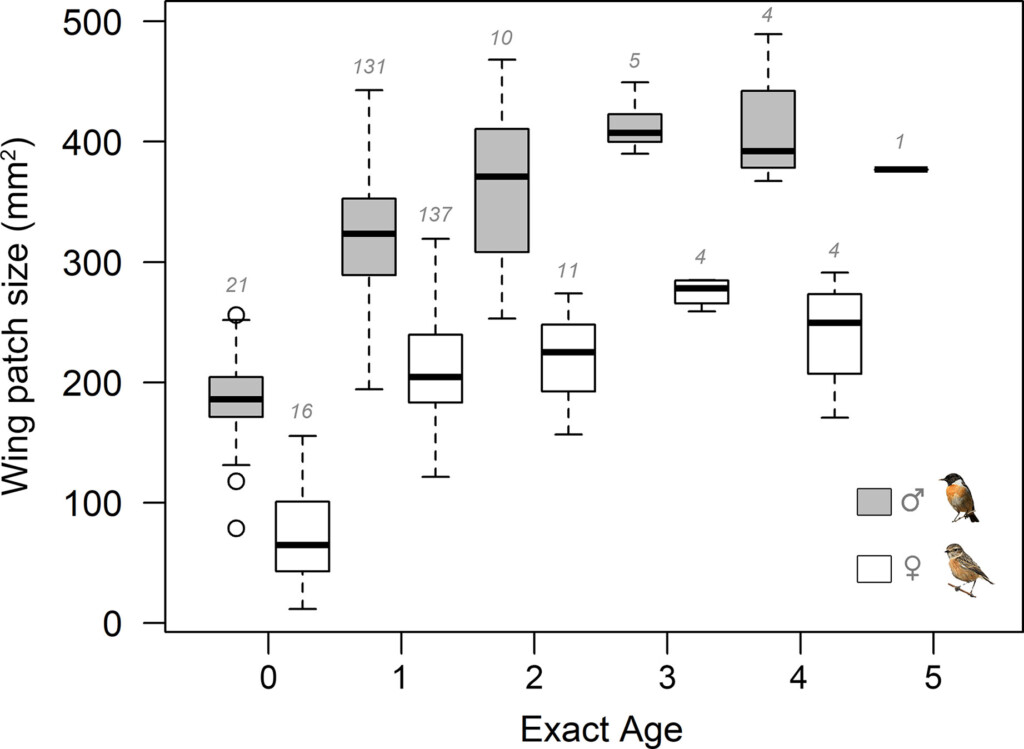
Figure 1. Wing patch size in male (dark grey) and female (white) Stonechats of known age. The horizontal bars indicate the median, the lower and upper limits indicate respectively the 1st and 3rd quartiles and the whiskers indicate the minimum and maximum values. Numbers above boxplots indicate sample sizes. Note that exact age 0 corresponds to juvenile plumage.
White wing patch variability
Overall, the findings suggest that there is substantial variability in the size of the white wing patch of male and female European Stonechats. The wing patch size varied both between and within individuals, with the largest source of variation at the population level being the sex and age of individuals.
The white wing patch was shown to be larger in males than in females for all age-classes. First-year individuals exhibited smaller wing patches than older birds, particularly males, and the researchers suggest that age-related increases in wing patch size at the population level might be mostly due to improvements within individuals. Males and older birds having larger patch sizes is consistent with other studies (e.g. Török et al. 2003).
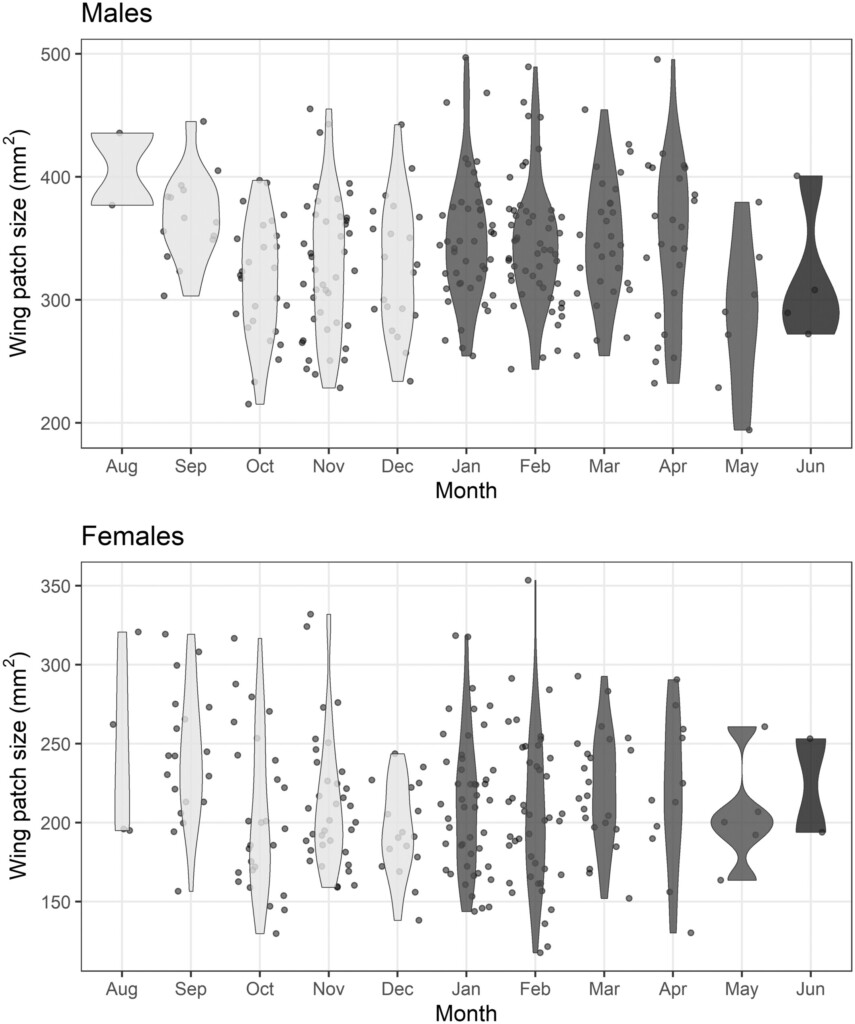
Figure 1. Variation in wing patch size throughout the plumage-year in male (top) and female (bottom) Stonechats. The kernel density plot shows the probability density of data (measurements), in each month. The grey scale represents the pre-breeding (light grey), breeding (mid-intensity grey) and moulting (dark grey) periods in our Stonechat population. Note that no birds were captured in July.
The size of the white wing patch varied between and within years. Other passerine studies have attributed seasonal changes in plumage ornament size to wear of feather tips and abrasion by preening and dust bathing (e.g. Møller & Erritzøe 1992). At the population level, Stonechat wing patch size decreased throughout the plumage-year, stabilising in the breeding season, which the researchers suggest could be due to its function as a potential signal during aggressive interactions and courtship displays. High quality individuals could develop more durable feathers (Fitzpatrick 1998) reducing wear of the melanin feathers surrounding the white patch, and/or invest more in preening (Delhey et al. 2006) reducing the soiling of the white feathers, affecting functioning of the trait.
The results also showed that when the mean temperature of the month prior to moult was higher, male birds tended to exhibit larger wing patches in the subsequent moult. Environmental factors can affect individuals and their ability to modify individual phenotypes, for example temperature and rainfall can impact food availability (Studds & Marra 2011), and high temperature may increase feather wear and tear (Scordato et al. 2012).
The results indicate that, although white wing patch size is a dynamic trait throughout the moults, there was substantial individual consistency in the trait throughout the life of individuals. Therefore, depigmented plumage ornaments may be indicators of long-term individual quality in spite of their phenotypic plasticity, reinforcing the idea that dynamic sexual traits could reflect genetic quality. However, further studies are needed to elucidate the exact function of the white wing patch in Stonechats, and determine whether the white patch acts as an independent signal or in conjunction with other plumage traits.
References
Delhey, K., Peters, A., Johnsen, A. & Kempenaers, B. (2006). Seasonal changes in blue tit crown color: do they signal individual quality. Behavioral Ecology 17: 790-798. VIEW
Fitzpatrick, S. (1998). Colour schemes for birds: structural coloration and signals of quality in feathers. Annales Zoologici Fennici 35: 67–77. VIEW
Møller, A.P. & Erritzøe, J. (1992). Acquisition of breeding coloration depends on badge size in male house sparrows Passer domesticus. Behavioral Ecology and Sociobiology 31: 271–277. VIEW
Scordato, E.S.C., Bontrager, A.L. & Price, T.D. (2012). Cross-generational effects of climate change on expression of a sexually selected traits. Current Biology 22: 78–82. VIEW
Studds, C.E. & Marra, P.P. (2011). Rainfall-induced changes in food availability modify the spring departure programme of a migratory birds. Proceedings of the Royal Society B: Biological Sciences 278: 3437–3443. VIEW
Svensson, L., Grant, P.J., Mullarney, K. & Zetterstrom, D. (1999). Collins Bird Guide. London, UK: Harper Collins Publishers.
Török, J., Hegyi, G. & Garamszegi, L.Z. (2003). Depigmented wing patch size is a condition-dependent indicator of viability in male collared flycatchers. Behavioral Ecology 14: 382-388. VIEW
Image credits
Top right: A female European Stonechat (Saxicola rubicola) | Alexis Lours | CC BY 4.0 Wikimedia Commons
Blog posts express the views of the individual author(s) and not those of the BOU.
If you want to write about your research in #theBOUblog, then please see here

